
Abioxénese
Este artigo trata sobre o estudo científico da orixe da vida. Para unha exploración máis ampla das diversas teorías relacionadas coa orixe da vida, incluíndo contextos históricos e filosóficos, véxase Teorías acerca da orixe da vida. Para os aspectos filosóficos e relixiosos véxase Cosmogonía. Para a evolución histórica anterior á refutación experimental da xeración espontánea véxase Teoría da xeración espontánea.

Nas ciencias naturais, a orixe da vida na Terra relaciónase coa teoría da abioxénese no sentido moderno que se lle dá en ciencia a este termo, e que estuda o como e o cando a vida apareceu e se desenvolveu na Terra e, hipoteticamente, noutros lugares do universo coñecido, a partir do big bang (datado hai uns 13.700 millóns de anos) até os nosos días.[1]
A cuestión da orixe da vida na Terra xerou nas ciencias da natureza un campo de estudo especializado cuxo obxectivo é dilucidar como e cando xurdiu. A orixe da vida na Terra, a partir da materia inerte, pódese datar, cos coñecementos actuais, nun período comprendido entre hai uns 4.400 millóns de anos, cando se deron as condicións para que o vapor de auga puidera condensarse por primeira vez, e permanecer en estado líquido na superficie terrestre, e hai uns 2.700 millóns de anos, cando apareceron os primeiros indicios de vida.[2]
A orixe da vida xa foi tratada na antigüidade no ámbito de diversas relixións e por diversas escolas filosóficas. O contraste entre os modelos científicos e o que se afirma literalmente nos textos sagrados das diversas relixións orixinou debates, ás veces virulentos, entre a ciencia e a fe.[3]
Desde o punto de vista científico, a explicación da orixe da vida parte do presuposto fundamental de que as primeiras formas viventes se orixinaron a partir de materia non vivente, a través de reaccións que actualmente non se producen no noso planeta. A interrogante sobre como se orixinou a vida sobre a Terra púidose comezar a responder despois do desenvolvemento da teoría da evolución por selección natural, elaborada por Alfred Russel Wallace e Charles Darwin en 1858. Esta teoría suxería que todas as formas de vida están relacionadas, por procederen dun antepasado común extremadamente "simple" desde o punto de vista biolóxico.
Pero o principal problema é comprender como se orixinou esta simple forma primordial, presumibelmente unha célula moi similar aos actuais procariotas, contendo a información xenética, conservada nos ácidos nucleicos, ademais de proteínas e outras biomoléculas indispensábeis para a propia supervivencia e reprodución. O proceso evolutivo que levou á formación dun sistema complexo e organizado (o primeiro ser vivente) a partir do mundo prebiótico durou seguramente centos de millóns de anos, a través de etapas sucesivas de fenómenos que, despois dun número elevado de tentativas, e grazas á intervención da selección natural, conduciron a sistemas progresivamente máis complexos. A primeira etapa fundamental foi a produción de moléculas orgánicas, como aminoácidos e nucleótidos, que constitúen "os ladrillos da vida" (o que se coñece como síntese química).
Os experimentos de Stanley Miller e outros demostraron que esta síntese era realizábel nas supostas condicións fisicoquímicas da Terra primordial, caracterizada por unha atmosfera redutora. Por outra parte, o achado de moléculas orgánicas no espazo, no interior de nebulosas, cometas e meteoritos, demostrou que estas reaccións se produciron tamén noutros lugares do universo, polo que algúns científicos supuxeron que as primeiras biomoléculas foron transportadas á Terra por medio de meteoritos.
A cuestión máis difícil é explicar como destes sinxelos compostos orgánicos, concentrados nos mares e constituíndo un chamado caldo primitivo, puideron chegar a formarse células dotadas dos requisitos mínimos esenciais para poderen ser consideradas viventes, isto é, a capacidade de utilizar materiais presentes no ambiente para manter a propia estrutura, organización, e poder reproducirse.
Moitos científicos trataron de aclarar, mediante hipóteses e experimentos, as etapas fundamentais que conduciron á vida, como a orixe dos primeiros polímeros biolóxicos e, despois disto, dunha molécula capaz de producir copias dos mesmos, o "replicador", do que derivan os nosos xenes, e a formación da primeira membrana biolóxica, que crearían compartimentos illados do ambiente externo, nos que evolucionaron os primeiros sistemas de reaccións e a primeira vía metabólica catalizada por enzimas. Non obstante, a reconstrución da historia da vida presenta aínda moitos interrogantes, concernentes sobre todo á sucesión dos eventos. Os progresos neste campo de investigación vense obstaculizados pola carencia de rexistros fósiles e pola dificultade de reproducir estes procesos no laboratorio.
Materia viva e materia inerte[editar | editar a fonte]
Que é a vida?[editar | editar a fonte]
- Véxase tamén: Vida.

Esta pregunta foi unha constante para os homes de todos os tempos, que intentaron atopar o porqué da diferenza entre os seres vivos e os que non o son. Aínda hoxe descoñecemos a natureza da vida, de xeito que non é posíbel dar unha definición concisa dela. Os biólogos cren que poden diferenciar "o vivente" do "non vivente", aínda que esta diferenciación, nalgúns casos, non sexa fácil.
Desde os primeiros momentos da historia do pensamento da nosa civilización occidental, as ideas sobre a vida encadráronse en dúas doutrinas filosóficas opostas: as materialistas ou mecanicistas, que supoñían que a vida non era máis que o resultado dunha organización máis ou menos complexa da materia, e as vitalistas ou finalistas, que propoñían que a vida tiña a súa orixe nunha forza superior que insuflaba aos seres un principio vital que, no caso dos homes, se identificaba coa alma.
Os primeiros defensores destas teorías foron dous filósofos da Grecia clásica, Demócrito (460-370 a. C.), influente filósofo presocrático, discípulo de Leucipo, famoso por formular unha teoría atómica para o cosmos, e Aristóteles (384 a. C.-322 a. C.), filósofo, lóxico e científico da Antiga Grecia, discípulo, desde os 17 anos, de Platón, co que estudou na súa famosa Academia en Atenas.[4][5]

O primeiro deles, claramente materialista, supoñía que toda a materia estaba formada por unhas diminutas partículas, tan pequenas que el mesmo as chamou átomos (grego ἄτομος, átomos, indivisible, de α-, 'non-' + τέμνω - temno, 'cortar'). Estes átomos, segundo Demócrito, serían os responsábeis das características da materia.
Así, por exemplo, o lume queimaba porque estaba formado por átomos cubertos de espiñas, do mesmo xeito que as propiedades da auga se debían a que os seus átomos eran moi lixeiros e posuían unha superficie lisa. Seguindo esta teoría, o filósofo grego supoñía entón que a vida se debía unicamente a que os seres que a posuían tiñan un tipo particular de átomos arredondados que, dispersos por todo o organismo, lles proporcionaban as características vitais.
Totalmente oposto a esta teoría, Aristóteles mantiña que os seres vivos estaban compostos de idénticos elementos que a materia inerte, pero que, a diferenza desta, posuían unha forza ou principio vital concedido por un ser superior. Este principio vital considerábase inmortal, non tendo a vida fin en si mesma, senón en función do seu Creador.
Aínda que ao longo dos anos ambas as teorías sufriron modificacións, a polémica sobre o materialismo e o vitalismo foi unha constante histórica influída máis por doutrinas filosóficas e relixiosas ca por un estrito pensamento científico.
Desde o punto de vista científico, as teorías materialistas e vitalistas pretenden definir un concepto abstracto de vida, mentres que o estudo dos seres vivos, da súa orixe e da súa evolución está baseado unicamente nos datos obxectivos sobre estes seres: a súa organización, a súa estrutura e o seu comportamento.[6]
A definición de morte[editar | editar a fonte]
Por outra parte, o concepto de vida ou existencia é inseparábel do de morte ou inexistencia, e a súa transcendencia, foi e é diferente nos distintos lugares e épocas da historia da humanidade. A importancia primordial da vida para o ser humano compróbase na súa influencia na linguaxe, de forma que son numerosos os diferentes usos e expresións que conteñen este termo. Só existe vida naquilo que pode morrer. Cada un dos seres vivos que existen sobre o noso planeta presentan un ciclo vital: nalgún momento nacen, durante un período máis ou menos longo desenvolven a súa vida e, finalmente, morren.
A discusión filosófica sobre a definición de vida vai unida á propia definición de morte, sobre todo cando esta se refire aos seres humanos. Así, para os filósofos vitalistas, a morte dunha persoa prodúcese no momento no que a alma abandona o corpo, mentres que os materialistas entenden a morte como unha fase máis do ciclo da materia.[7]
As funcións vitais[editar | editar a fonte]
Se dar unha definición en abstracto do que é a vida supón unha tarefa difícil, diferenciar entre un ser vivo e outro que non o é parece relativamente simple. Para iso debemos atopar unha forma de describir o que queremos dicir cando dicimos que imos estudar algo que está vivo. E facémolo recoñecendo certos trazos, certas características, que son comúns a todos os seres vivos e non o son aos non viventes.



- Complexidade. As rochas están formadas, como moito, por uns poucos minerais esenciais e outros poucos accesorios, é dicir, por unhas poucas moléculas diferentes. En cambio os seres vivos, incluso a máis simple das bacterias, están formados por centos de miles de moléculas distintas, e as máis delas, ademais, extraordinariamente complexas.
- Organización. Podemos distinguir na materia diferentes niveis de organización. Para que poida constituírse un ser vivo, o nivel mínimo de organización é o de organización celular: todos os organismos están constituídos por unha ou por moitas células.
- Crecemento e diferenciación. Para crecer e para manter a súa organización corporal, os organismos necesitan achegas de materia e de enerxía, que toman do exterior e transforman en materia propia e na enerxía necesaria para o mantemento da actividade vital, devolvendo ao medio externo as materias que non lles serven, e tamén parte da enerxía. Estes intercambios de materia e enerxía constitúen unha serie de funcións denominadas no seu conxunto nutrición. O conxunto de todas as reaccións químicas que se producen no organismo reciben o nome de metabolismo, que comprende dúas fases: unha que incorpora as substancias coas que o ser vivo constrúe as súas propias moléculas, chamada anabolismo ou asimilación, e outra, o catabolismo ou desasimilación, na que se destrúen parte dos produtos obtidos na fase anterior, empregando a enerxía liberada para manter todas as funcións.
- Irritabilidade ou excitabilidade. É a capacidade que teñen de reaccionaren fronte aos estímulos. Os estímulos son calquera cambio físico ou químico que se produza no exterior ou no interior do organismo e que pode ser detectado por este. Grazas a esta propiedade, os seres vivos son capaces de recoñecer o medio no que viven e de responder aos cambios que nel se producen. Un tipo especial de resposta é a adaptación dos organismos aos cambios operados no medio, que constitúe un dos principais aspectos da evolución biolóxica.
- Autorregulación. É o control que todo ser vivo exerce sobre as súas funcións mediante unha serie de reaccións químicas que se realizan en diferentes órganos e que lle permiten ao ser vivo adaptarse e relacionarse co medio que o rodea (e que por iso chamamos funcións de relación), tanto no que respecta ao ambiente físico como ao resto de seres vivos que habitan con el. Grazas a estas funcións pode buscar seus alimentos, defenderse dos seus inimigos, buscar outros individuos de sexo oposto (no caso da reprodución sexual), adaptarse ás variacións do medio (e mesmo migrar a outras zonas máis apropiadas durante as estacións desfavorábeis) e, en fin, permanecer con vida no lugar en que habita.
- Reprodución ou autoperpetuación. Consiste na creación, por parte dos organismos, doutros organismos semellantes (non idénticos), que aseguran a permanencia das diferentes especies no planeta.
En resumo, os seres vivos teñen unha organización celular do seu corpo e realizan unha serie de funcións, chamadas funcións vitais por presentarse só nos seres vivos. Estas pódense reunir en tres grandes grupos ou tipos: de nutrición ou autoconservación, que serven para o mantemento do individuo; de relación ou autorregulación, que permiten ao individuo o recoñecemento do medio externo e interno, necesario para a nutrición e a reprodución; e as de reprodución ou autoperpetuación, que aseguran o mantemento da especie.[8]
Niveis de organización dos seres vivos[editar | editar a fonte]





O desenvolvemento de todas as ramas da bioloxía en xeral, e da bioquímica e a bioloxía celular en particular, permitiu estudar con todo detemento os seres vivos até as súas estruturas máis sinxelas. Así, púidose comprobar que todos eles están formados pola coordinación dunha serie de sistemas cada vez máis simples.
Ao estudarmos un animal superior podemos observar que está formado por unha serie de aparellos e sistemas (circulatorio, dixestivo, nervioso etc.) que, coordinados entre si, permiten a existencia do animal. Pero, á súa vez, estes aparellos están constituídos por órganos (corazón, pulmóns, estómago, encéfalo etc.), formando unha estrutura que dá lugar a cada un dos aparellos ou sistemas.
Este resultado, que podería continuar até estruturas cada vez menores, máis os datos obtidos por físicos e químicos, permiten afirmar que toda a materia existente na Natureza está dividida nunha serie de chanzos xerárquicos, de tal xeito que os elementos de cada un deles están formados pola estrutura organizada dun conxunto, maior ou menor, de elementos do nivel anterior. Esta escala de niveis de organización está constituída, de menor a maior grao de complexidade, por partículas elementais, átomos, moléculas, orgánulos, células, tecidos, órganos, sistemas de órganos ou aparellos, organismos, poboacións, comunidades, ecosistemas, biomas e biosfera, entendido este último termo como o conxunto de todos os seres vivos que existen na Natureza máis as condicións ambientais que os arrodean. Para facilitar o seu estudo, podemos reunir estes niveis en cinco grandes grupos: nivel molecular, nivel celular, nivel orgánico, nivel poboacional e nivel de ecosistema.
- Nivel molecular. Constitúeno as partículas subatómicas, ou elementais (protóns, neutróns e electróns) que, ao reunírense, forman os átomos. As moléculas están formadas pola unión, mediante enlaces químicos, de átomos iguais ou diferentes. As moléculas que forman parte dos seres vivos denomínanse biomoléculas. Moitas das biomoléculas son macromoléculas, é dicir, moléculas de gran tamaño. As macromoléculas son o resultado da unión de distintas moléculas, como é o caso dalgúns aminoácidos e nucleósidos, que dan lugar ás proteínas e aos ácidos nucleicos, respectivamente. A unión de varias macromoléculas pode dar lugar a asociacións macromoleculares, como ocorre nos complexos multienzimáticos, nas membranas celulares ou nos sistemas contráctiles das células. Estas, á súa vez, poden asociarse formando orgánulos, como as mitocondrias e os cloroplastos.
- Nivel celular. É o primeiro nivel que se pode considerar que inclúe seres vivos. A el pertencen as células procarióticas, que non posúen núcleo diferenciado, como as das bacterias, e as células eucarióticas, que posúen núcleo diferenciado e un citoplasma moi complexo: son as células dos animais, dos vexetais, dos fungos, das algas e dos protozoos. Os virus tamén están incluídos neste nivel para algúns autores, mentres que outros os sitúan no nivel anterior (como asociacións macromoleculares.
- Nivel orgánico. Inclúe todos os organismos, tanto unicelulares coma pluricelulares. Porén, nestes últimos ten lugar unha división do traballo fisiolóxico e, consecuentemente, unha diferenciación celular que orixina tecidos (como o muscular ou o nervioso). Os tecidos reúnense para formar órganos (como o corazón ou os pulmóns), cada un dos cales realiza unha función específica (denominada acto). O conxunto de todos os órganos que de forma coordinada desempeñan un traballo fisiolóxico de nivel superior (función) constitúe un aparello (como o dixestivo, ou o circulatorio).
- Nivel de poboación. Os seres vivos non están illados, senón que se relacionan entre eles. A relación máis importante é o emparellamento, que dá lugar á formación de novos individuos. Tamén adoitan producirse situacións de competencia polo alimento ou polo espazo, e situacións de protección mutua. Unha poboación é o conxunto dos individuos da mesma especie que viven nunha mesma zona e se inflúen mutuamente.
- Nivel de ecosistema. As distintas poboacións que habitan na mesma zona forman unha comunidade ou biocenose. As condicións fisicoquímicas da mesma forman o biótopo. O conxunto formado pola biocenose, o biótopo e as relacións que entre eles se establecen forman o ecosistema. Os factores climáticos delimitan zonas de vexetación típica que, á súa vez, condicionan a existencia nelas dunha fauna concreta, repetíndose ditas zonas en áreas moi extensas da Terra que reciben o nome de biomas. E o conxunto de todos os biomas do planeta constitúe a biosfera.[9][10][11][12]
En cada un destes niveis de organización danse dúas calidades: unidade e diversidade. Por exemplo, existe unha certa unidade nas clases de átomos e moléculas que compoñen todos os seres vivos; hai unha unidade na forma na que se agrupan as moléculas nos diferentes órganos etc. Porén, existe tamén unha gran diversidade pois, aínda estando formados por átomos iguais ou tendo estruturas básicas moi similares, cada un dos elementos encadrados nun nivel de organización posúen características diferenciais, distintos comportamentos —tanto a nivel físico coma químico— e, en fin, propiedades varias que permiten distinguir con exactitude cada un dos elementos dos outros.
A análise dos elementos dos distintos niveis de organización permite chegar, dentro da Natureza, a unha exacta diferenciación entre a materia viva e a inerte. Se observamos cada un destes niveis notaremos que é a partir dun deles, o dos orgánulos, cando aparecen as funcións vitais características dos seres vivos. Isto significa que todo nivel organizativo que se ache por debaixo deste, haberá de posuír o carácter de materia inerte, mentres que, a partir do nivel de orgánulo, toda a materia existente no universo será considerada como materia viva.
Os orgánulos son considerados o primeiro nivel de organización que empeza a presentar mostras de vida porque na súa composición química hai xa moléculas inexistentes na materia inerte —as biomoléculas— que serán as responsábeis das características propias de todo ser vivo.[6]
Á busca da orixe da vida[editar | editar a fonte]
A orixe da vida. Teoría da xeración espontánea[editar | editar a fonte]
- Artigo principal: Teoría da xeración espontánea.



Aínda que a exacta complexidade bioquímica e celular dos seres vivos non foi coñecida até hai uns poucos anos, sabíase desde sempre que a estrutura de animais e plantas era moito máis complexa cá da materia inerte. Hai xa moitos séculos que este coñecemento levou aos homes a preguntárense a orixe desta complexidade ou, o que é o mesmo, a orixe da vida. Se nos preguntamos hoxe pola orixe dun cabalo ou dun piñeiro, é fácil responder que estes proceden doutros seres vivos semellantes a eles. É fácil xeneralizar a resposta e concluír que a vida ten como orixe outra vida preexistente.
Pero non sempre foi así. A pouca base científica existente na antigüidade deu lugar a que a solución á pregunta da orixe da vida se achase en doutrinas máis filosóficas ca científicas, entre as que destacou a teoría da xeración espontánea, formulada inicialmente por Aristóteles. Aristóteles admitía que, en xeral, os seres vivos se orixinan doutros seres vivos semellantes, pero que igualmente podían orixinarse da materia inerte e, dado o enorme prestixio deste filósofo da antiga Grecia, tivo vixencia até mediado o século XIX, sobre todo porque a súa hipótese vitalista cadraba moi ben coas concepcións relixiosas do cristianismo. Esta tendencia representárona sobre todo os filósofos chamados neoplatónicos, á fronte dos cales estaba Plotino, quen consideraba a xeración espontánea como resultado da entrada na materia dun espírito vivificador.
Todo o pensamento da Idade Media estivo grandemente influído por Aristóteles e, por tanto, a crenza na xeración espontánea mantívose. Nesta época os chamados Pais da Igrexa, como San Basilio (igrexa oriental) ou Santo Agostiño (igrexa occidental), consideraban a xeración espontánea como unha verdade indiscutíbel, e moi de acordo coa Biblia, sempre que se aceptara que o espírito vivificador da materia era insuflado por Deus. Así mesmo defenderon estas ideas San Tomé e os escolásticos.
Mesmo no Renacemento, época na que a observación dos fenómenos naturais é cada vez maior e máis exacta, e na Idade Moderna, seguiuse a admitir esta teoría. Personaxes como William Harvey (1578-1657), célebre polo descubrimento da chamada circulación maior do sangue, pese a pronunciar a famosa frase omne vivum ex ovo (todo ser vivo procede dun ovo), segue admitindo a posibilidade da xeración espontánea para animais inferiores. No mesmo caso están Jan Baptista van Helmont (1577-1644), autor de numerosos traballos sobre fisioloxía vexetal, René Descartes (1596-1650) ou Isaac Newton (1642-1727), que estableceron leis exactas da natureza pero aceptaron sen crítica ningunha a xeración espontánea.[13]
O primeiro en enfrontarse ao dogma da xeración espontánea foi o italiano Francesco Redi (1626-1697) que cun sinxelo experimento, consistente en deixar durante varios días uns anacos de carne en frascos, parte deles descubertos e parte pechados hermeticamente, comprobou que na carne situada nos frascos abertos apareceran gran número de vermes, e moscas voando, pero nos frascos pechados non aparecía ningún. Os seus detractores consideraron erróneo o experimento, xa que se non aparecían vermes nos frascos pechados era debido á falta de aireación. Redi repetiu o experimento cun terceiro grupo de frascos, pechados cunha tea porosa, que deixaba pasar o aire pero non ás moscas, e tampouco nestes apareceron vermes. Pero nin con esas. Ademais, a influencia da xeración espontánea era tan grande, que o mesmo Redi admitíaa nalgúns casos, coma nos vermes intestinais ou nos da madeira (larvas de couzas).
Algo máis tarde, e debido aos descubrimentos dos animálculos (microorganismos) polo holandés Anton van Leeuwenhoek (1632-1723), biólogo afeccionado e excepcional observador, deron novo pulo ás discusións e volveron a repetirse experimentos que permitiran comprobar a veracidade ou falsidade da xeración espontánea, aínda que agora limitadas aos seres microscópicos. A polémica de máis sona foi a que mantiveron o británico John Needham (1713-1781) e o italiano Lazzaro Spallanzani (1729-1799). Ambos os dous cultivaron microorganismos en caldos nutritivos quentando os recipientes, pechados hermeticamente, que os contiñan, aparecendo nos de Needham microorganismos. Spallanzani replicou dicindo que o inglés non quentaba suficientemente o caldo, como facía el, que o chegaba a facer ferver (é dicir, o esterilizaba, que diriamos hoxe) e, daquela, os microorganismos no aparecían. Needham retrucou dicindo que "a calor excesiva" destruía o principio vital. A maioría dos científicos contemporáneos déronlle a razón a Needham.
Xa no século XIX, concretamente en 1836, o fisiólogo alemán Theodor Schwann (1810-1882), famoso pola súa contribución ao establecemento da teoría celular, introduciu nos caldos de cultivo aire esterilizado por quecemento, non aparecendo nos cultivos microbio ningún. Pero a pesar desta evidencia, moitos científicos da época aínda mantiñan que a esterilización (aínda que fora a do aire, e non do caldo) destruía a forza vital. Ademais, o progresivo coñecemento de microorganismos como as bacterias favoreceu o mantemento da idea espontaneísta.[14]
A Academia de Ciencias de Francia, por aquelas datas, ofrecera un premio ao investigador que lograra solucionar definitivamente tan vello problema. Dito premio concedéuselle ao físico e químico, pero sobre todo bacteriólogo, Louis Pasteur (1822-1895), en 1862. O mérito de Pasteur foi enorme, xa que foi quen de probar a invalidez dunha teoría que se mantivera durante vinte e cinco séculos. O sabio francés comezou por aclarar a presenza de microorganismos vivos no aire, para o que ideou un mecanismo de aspiración do mesmo a través dun tubo obturado con nitrato de celulosa; despois de vinte e catro horas extraía a nitrocelulosa, na que se observaba un sedimento que, lavado e observado ao microscopio, demostraba que alí había, efectivamente, microorganismos.
Para demostrar que estes microorganismos do aire estaban vivos e eran a causa da descomposición dos líquidos orgánicos, e non o resultado da mesma, introduciu líquidos orgánicos en recipientes de fondo redondo cos colos estirados e unidos a un tubo de platino que ía quentando até poñelo incandescente; despois arrefriaba o aire mediante unha corrente de auga antes de que penetrara no recipiente e, unha vez cheo este de aire, pechaba o colo á chama dun queimador de gas. Nestas condicións, os líquidos mantíñanse inalterábeis. Quedaba, pois, demostrado que os líquidos orgánicos postos en contacto co aire estéril non se alteraban, nin tampouco se producían no seu seo xéneses espontáneas de organismos vivos, e que calquera destes fenómenos dependía exclusivamente dos microorganismos do aire, sendo por tanto desbotada para sempre a teoría da xeración espontánea.[15]
- Darwin. Nunha carta a Joseph Dalton Hooker o 1 de febreiro de 1871, Charles Darwin escribiu:
"Dise a miúdo que hoxe en día se dan todas as condicións para a produción dun organismo vivo, e que ditas condicións puideran existir sempre. Pero se puideramos concibir que nalgunha poza cálida, na que houbera toda clase de sales fosfóricos e de amonio, luz, calor, electricidade etc., que un composto proteico se formara por medios químicos e listo para sufrir cambios aínda máis complexos, a día de hoxe ese tipo de materia sería instantaneamente destruído ou absorbido, o que non sucedería no caso de que os seres vivos aínda non apareceran."[16]
Noutras palabras, a presenza da vida mesma fai a busca da orixe da vida dependente das condicións de esterilidade que se dan no laboratorio. Máis precisamente, o osíxeno producido polas diferentes formas de vida é moi activo a escala molecular, o cal prexudica calquera intento de formación de vida.
- A evolución e a súa relación coa orixe da vida. Discutiuse se a orixe da vida e a orixe do proceso de evolución xurdiron ao mesmo tempo. Postulouse que, de forma equivalente a como actúa o proceso de evolución nos seres vivos, tamén actuarían os mecanismos evolutivos en compostos químicos antes de que houbera vida. Neste sentido, científicos como Martin A. Nowak e Hisashi Ohtsuki postularon o como e o cando a cinética química pasa a converterse nunha dinámica evolutiva, formulando unha teoría matemática xeral para a orixe da evolución. Nela descríbese a previda coma un alfabeto de monómeros que forman ao azar polímeros, sendo un sistema xerativo que pode producir a información, na que orixinalmente se presenta unha preevolutiva dinámica de selección e mutación, pero non replicación, a diferenza da vida. A partir de análise matemático conclúese que as mellores e máis competentes candidatas moleculares para a vida xa foran seleccionadas antes incluso de que empezaran a reproducirse. Igualmente, existe unha fase de transición na que, se a taxa efectiva de replicación supera un valor crítico, entón a vida compite coa previda e, finalmente, a vida destrúe á previda.[17]
A cuestión da orixe da vida: condicións ambientais na Terra primixenia[editar | editar a fonte]
Os fundamentos[editar | editar a fonte]

_mathilde.jpg)
Unha vez sentada, grazas a Pasteur, a base de que a vida procede de si mesma, quedaba por aclarar o como e o cando se orixinou a primeira vida sobre a Terra. Para tratar de esclarecer este punto déronse dúas circunstancias particularmente favorábeis: a formulación da teoría da evolución por Charles Darwin (1809-1882), e o enorme desenvolvemento da física, a química e a xenética. Os datos achegados pola teoría evolucionista e as disciplinas anteriormente citadas suscitan o problema da orixe da vida dentro dun contexto evolutivo.
A idea de utilizar a evolución para explicar o fenómeno vital non é nova. O francés Jean-Baptiste de Monet, coñecido por Lamarck (1744-1829) suxerira que a materia mineral podería xerar substancias orgánicas que poderían ordenarse pouco a pouco. Non obstante, habería que agardar aínda bastantes anos para que se formularan teorías máis completas.
Realmente, durante o século XIX sentaron as bases de todo o coñecemento biolóxico posterior. Os fitos máis salientábeis, por orde cronolóxica, foron os seguintes:
- 1828: O químico alemán Friedrich Wöhler sintetiza a urea (molécula orgánica) a partir de dióxido de carbono e amoníaco (moléculas minerais), rompendo así a barreira que até entón existía entre a materia orgánica e a materia mineral: a materia é única.
- 1834: Establécese a teoría celular: todos os seres vivos están formados por células.
- 1858: Wirchow postula que toda célula procede doutra célula preexistente: Omnis cellula ex cellula.
- 1862: Pasteur desbota definitivamente a teoría da xeración espontánea: toda vida procede doutra anterior.
- 1859: Darwin, non sen polémicas, dá resposta ao problema da orixe das especies mediante a súa teoría da evolución por selección natural.
- 1860: Mendel constrúe o modelo básico que interpreta a herdanza biolóxica (aínda que os seus experimentos e conclusións non se coñeceron até o inicio do século XX).
Primeiros pasos[editar | editar a fonte]
Pero non puido ser até o século XX, grazas ao progresivo coñecemento da composición química dos seres vivos, por unha parte, e da orixe e evolución do sistema solar e do planeta Terra, por outra, cando se puido abordar cientificamente o problema da orixe da vida sobre o noso planeta.
O corpus de estudos sobre a orixe da vida forma unha área limitada de investigación, a pesar do seu profundo impacto na bioloxía e a comprensión humana do mundo natural. Co obxectivo de reconstruír o evento empréganse diversos enfoques baseados en estudos tanto de campo como de laboratorio. Por unha parte o ensaio químico no laboratorio e, por outra, a observación de procesos xeoquímicos ou astroquímicos que produzan os constituíntes da vida nas condicións nas que se pensa que puideron suceder no seu contorno natural. Na tarefa de determinar estas condicións tómanse datos da xeoloxía da idade escura da Terra a partir de análises radiométricas de rochas antigas, meteoritos, asteroides, así como a observación astronómica de procesos de formación estelar. Por outra parte, inténtanse achar as pegadas presentes nos actuais seres vivos daqueles procesos mediante a xenómica comparada e a busca do xenoma mínimo. E, por último, trátase de verificar as pegadas da presenza da vida nas rochas, como microfósiles, desviacións na proporción de isótopos de orixe bioxénica e a análise de contornas, moitas veces extremófilas, semellantes aos paleoecosistemas iniciais. Existen unha serie de observacións que intentan describir as condicións fisicoquímicas nas que puido emerxer a vida, pero aínda non se ten un cadro razoabelmente completo acerca de como puido ser esta orixe. Propuxéronse varias teorías, sendo a hipótese do mundo de ARN e a teoría do mundo de ferro-xofre as máis consideradas pola comunidade científica.[18]
Para o coñecemento das condicións iniciais emprégase a teoría xeoquímica no estudo das rochas antigas, efectúanse simulacións de laboratorio e por medio de ordenadores. Un dos puntos centrais é determinar a dispoñibilidade de elementos e moléculas esenciais, en especial metais, posto que son indispensábeis como cofactores na bioquímica actual, así como o seu estado redox nas distintas localizacións.[19] Así mesmo, é esencial datar as primeiras manifestacións da vida para aproximar o lapso de tempo no que estamos buscando.
Composición da atmosfera, os océanos e a codia terrestre no eón Hadeico[editar | editar a fonte]
A formación e acreción da Terra tivo que ter lugar, ao mesmo tempo que o sistema solar, hai uns 4.600 millóns de anos, como resultado da condensación de po cósmico e gases interestelares, e de sucesivos choques de meteoritos, segundo diferentes métodos radiométricos. Nos primeiros tempos sería unha especie de bóla quente na que a superficie (e as zonas máis internas) estaban fundidas. A medida que foi arrefriando formouse a codia sólida, constituída principalmente por silicatos. As rochas máis antigas coñecidas, situadas en Groenlandia, datan de hai uns 3.800 millóns de anos. Hai suficientes indicios nestas e noutras rochas antigas como para poder reconstruír a historia da Terra nos seus primeiros momentos, e saber que tipo de ambiente predominaba na súa superficie.
A diferenciación do manto terrestre, a partir de análises da serie samario/neodimio en rochas de Isua, Groenlandia, puido ser bastante rápida, talvez en menos de 100 millóns de anos.[20] Posteriores estudos confirman esta formación temperá das capas de silicatos terrestres.[21]
Ao mesmo tempo que a codia terrestre formouse a atmosfera primitiva, de carácter máis ou menos redutor, composta por vapor de auga, dióxido de carbono, metano, amoníaco, anhídrido sulfuroso e cloruro de hidróxeno. Non había osíxeno libre. Ao descender a temperatura, o vapor de auga da atmosfera condensou e caeu en forma de chuvia, o que terminou por formar os océanos, ao acelerarse o arrefriamento da superficie e permitir a existencia de auga líquida.
Suxeriuse que os océanos poderían aparecer no eón Hadeico tan cedo como hai 200 millóns de anos despois da formación da Terra,[22] nun ambiente quente (100 ̊C) e redutor, e cun pH inicial de 5,8 que subiu rapidamente cara á neutralidade. Esta idea foi apoiada por Wilde[2] quen elevou a datación dos cristais de circón encontrados en cuarcitas metamorfizadas do terrane do gneis de Monte Narryer, en Australia occidental, do que previamente se pensaba que era de 4.100 - 4.200 millóns de anos a 4.402 millóns de anos. Outros estudos realizados máis recentemente no cinto de basalto de Nuvvuagittuq, ao norte do Quebec, empregando neodimio-142 confirman, estudando rochas do tipo falsa anfibolita (cummingtonita-anfibolita), a existencia moi temperá dunha codia, cunha datación de 4.360 millóns de anos.[23]
Isto significa que os océanos e a codia continental existiron dentro dos 150 primeiros millóns de anos despois da formación da Terra. A pesar disto, o ambiente hadeico era enormemente hostil para a vida. Serían frecuentes as colisións con grandes obxectos cósmicos, incluso de máis de 500 quilómetros de diámetro, suficientes para vaporizar o océano durante meses tras o impacto, o que formaría nubes de vapor de auga mesturado con po de rochas ascendendo a elevadas altitudes que cubrirían todo o planeta. Despois duns cantos meses a altitude desas nubes comezaría a diminuír, pero a base da nube continuaría aínda estando elevada probabelmente durante os seguintes mil anos, despois do cal comezaría a chover a unha altitude máis baixa. Durante 2.000 anos as chuvias consumirían lentamente as nubes, devolvendo os océanos á súa profundidade orixinal só 3.000 anos despois do impacto.[24]
O posíbel bombardeo intenso tardío provocado probabelmente polos movementos posicionais dos planetas gasosos xigantes, que cribaron a Lúa e os planetas interiores (Mercurio, Marte e, posibelmente, a Terra e Venus) hai de 3.800 a 4.100 millóns de anos probabelmente esterilizarían o planeta se a vida xa aparecera nese período.
Primeiras evidencias directas da aparición da vida[editar | editar a fonte]

Segundo as evidencias actuais, aínda que están suxeitas a controversia, a vida debeu aparecer tras o arrefriamento do planeta que seguiu ao bombardeo intenso tardío, hai uns 4.000 millóns de anos. Aínda que todos os seres vivos actuais parecen provir dun único organismo ancestral, neste asunto cabe preguntarse se houbo varias aparicións "fortuítas" de formas de vida das que só sobreviviu unha, ou se ben esas formas de vida aínda sobreviven porque non sabemos buscalas.[19]
Unha das formas de verificar a actividade biolóxica é unha curiosa propiedade dos sistemas celulares, como a fotosíntese, que incorporan CO2 de diversas fontes para transformalo en moléculas orgánicas. Existen dous isótopos estábeis do carbono, C12 e C13, permanecendo estábeis as súas abundancias relativas na atmosfera. Cando se incorpora o CO2 por un sistema biolóxico, este prefire o isótopo máis lixeiro, arrequecendo as rochas carbonatadas no outro isótopo.[25]
A proba dunha aparición temperá da vida vén do cinto supracortical de Isua en Groenlandia occidental e de formacións similares nas veciñas illas de Akilia. O carbono que forma parte das formacións rochosas ten unha concentración de 13C elemental de aproximadamente -5,5, o que debido a que en ambiente biótico adoita preferirse o isótopo máis lixeiro do carbono,12C, a biomasa ten unha 13C de entre -20 e -30. Estas "sinaturas" isotópicas consérvanse nos sedimentos e Mojzis[25] usou esta técnica para suxerir que a vida xa existiu no planeta hai 3.850 millóns de anos. Lazcano e Miller (1994) suxiren que a rapidez da evolución da vida está determinada pola taxa de auga recirculante a través das fumarolas submarinas centrooceánicas. A recirculación completa leva uns 10 millóns de anos, por iso calquera composto orgánico producido daquela podería alterarse ou destruírse por temperaturas que excedan os 300 ̊C. Ambos estiman que o desenvolvemento a partir dun xenoma de 100 quilobases dun heterótrofo primitivo de ADN/proteínas até a xeración dun xenoma de 7.000 xenes dunha cianobacteria filamentosa requiriría tan só 7 millóns de anos.[26]
Bioxénese en ambiente cálido contra frío[editar | editar a fonte]
Se se examinan os períodos libres de cataclismos producidos por impactos de meteoritos que impedirían o establecemento de protoorganismos autorreplicantes, a vida puido desenvolverse en diferentes ambientes primitivos. O estudo levado a cabo por Maher e Stephenson[27] amosa que se os sistemas hidrotermais mariños profundos propician un lugar aceptábel para a orixe da vida, e a abioxénese puido suceder en datas tan temperás como entre hai 4.000 e hai 4.200 millóns de anos, mentres que se sucedera na superficie da Terra a abioxénese só podería ocorrer hai entre 3.700 e 4.000 millóns de anos.
Outros traballos de investigación suxiren un comezo da vida máis frío. Os traballos de Stanley Miller mostraron que os ingredientes da vida adenina e guanina requiren condicións de conxelación para a súa síntese, mentres que a citosina e o uracilo precisan temperaturas de ebulición.[28] Baseándose nestas investigacións suxeriu que a orixe da vida implicaría condicións de conxelación e meteoritos impactando.[29]
Un artigo da publicación Discover Magazine apunta cara á investigación de Stanley Miller indicando que se poden formar sete aminoácidos diferentes e 11 tipos de nucleobases en xeo, como cando se deixou amoníaco e cianuro no xeo antártico entre 1972 e 1997,[30] así como á investigación levada a cabo por Hauke Trinks amosando a formación de moléculas de ARN de 400 bases de lonxitude en condicións de conxelación utilizando un molde de ARN (unha cadea sinxela de ARN que guía a formación dunha nova cadea). A medida que a nova cadea de ARN medraba, os novos nucleótidos íanse adherindo ao molde.[31]
A explicación que se deu para a inusitada velocidade destas reaccións a semellante temperatura é que se trataba dunha conxelación eutéctica. A medida que se forman cristais de xeo, este permanece puro: só se unen as moléculas de auga ao cristal en crecemento, mentres que as impurezas como o sal ou o cianuro quedan excluídas. Estas impurezas acaban amoreadas en bolsas microscópicas de líquido entre o xeo, e é esa concentración o que fai que as moléculas choquen entre si con máis frecuencia.[32]
A síntese abiótica[editar | editar a fonte]
Oparin e Haldane[editar | editar a fonte]
- Artigos principais: Aleksandr Ivánovich Oparin e John Burdon Sanderson Haldane.
Unha vez desbotada a xeración espontánea, a cuestión da orixe da vida centrábase en intentar explicar a orixe da primeira célula. Os coñecementos da astronomía e a orixe do sistema solar permitían especular sobre as condicións en que xurdira este sistema vivo. Até 1924 non se realizou ningún progreso real, cando o bioquímico ruso Aleksandr Oparin (1894-1980) demostrou experimentalmente que o osíxeno atmosférico impedía a síntese de moléculas orgánicas, que son os constituíntes necesarios para o xurdimento da vida. Nese mesmo ano publicou unha pequena obra titulada A orixe da vida, onde expoñía unha teoría quimiosintética, segundo a cal as moléculas orgánicas puideran evolucionar fóra de todo organismo, reunirse e formar sistemas sometidos aos principios da evolución.

Por outra parte, en 1929, o pintoresco científico escocés John Burdon Sanderson Haldane (1892-1964), xenetista, biólogo evolutivo e un dos fundadores da xenética de poboacións, entre outras cousas, publicou un libro sobre a orixe da vida no que expoñía as súas ideas, semellantes ás de Oparin, e ás que chegou de forma independente. Hai que sinalar que o libriño de Oparin foi traducido ao inglés en 1939, polo que até esta data permaneceu practicamente descoñecido en occidente.
Na súa obra A orixe e o desenvolvemento da vida[33][34], Oparin desenvolvía a teoría quimiosintética. Para el as condicións existentes cando se formou a Terra serían totalmente diferentes ás que existen na actualidade, tratándose dunha atmosfera redutora que favorecería a formación de polímeros e dificultaría a súa hidrólise unha vez formados. Nestas condicións formaríanse moléculas orgánicas sinxelas a partir das cales se poden formar os compoñentes das proteínas, dos ácidos nucleicos) e dos azucres sinxelos. Estes compostos disolveríanse nos océanos, que se irían arrequecendo progresivamente en materia orgánica, formando o chamado caldo primitivo onde, segundo Oparin, xurdiría a primeira célula viva de modo espontáneo. As moléculas orgánicas iríanse asociando para formaren as macromoléculas que integran a materia viva. Este proceso iría acompañado dunha selección natural en virtude da cal unicamente perdurarían os coacervados que tiveran capacidade de autosíntese e de autoduplicación, que irían evolucionando cara a formas cada vez máis complexas e estábeis. Atopariámonos así nos albores do tipo de célula máis sinxela, a primeira forma de vida ou nivel celular.[6][35]
O mesmo ano Haldane tamén suxeriu que os océanos prebióticos da Terra, moi diferentes dos seus correspondentes actuais, formaran un "caldo quente diluído" no cal se poderían formar os compostos orgánicos. A esta idea chamouna biopoese (do grego βίος, "bíos", 'vida' e ποιήσις "poiésis", 'formación'), é dicir, o proceso polo cal a materia viva xorde de moléculas autorreplicantes pero non vivas.[36]
Non sabemos cando e como estes grupos moleculares grandes e complexos se perfeccionaron até convertérense en verdadeiras estruturas viventes. Probabelmente os agregados moleculares obterían a enerxía necesaria doutras moléculas orgánicas existentes no caldo primitivo e de procesos elementais de fermentación. Resultado destes últimos sería a acumulación cada vez maior de dióxido de carbono, creando talvez as condicións favorábeis para a aparición do proceso fotosintético.
O experimento de Miller[editar | editar a fonte]
- Artigos principais: Stanley Miller, Harold Clayton Urey e Experimento de Miller e Urey.

|

| |
A teoría de Oparin non deixa de ser unha hipótese. Pese a iso puido ser confirmada, en parte, grazas ao químico estadounidense Stanley Miller (1930-2007).
En 1953 era un estudante bolseiro, diplomado en química na Universidade de Chicago, alumno de Harold Clayton Urey (que fora premio Nobel de Química en 1934) e tivo a idea de simular nun balón de vidro a atmosfera primitiva coa mestura de gases proposta por Oparin e Haldane e bombardeala con descargas eléctricas de 60.000 volts, que simularían as radiacións solares ás que nesa época debería estar sometida a superficie do noso planeta.
Cando, ao cabo dunha semana, analizou os produtos resultantes das reaccións, comprobou que se sintetizaran compostos orgánicos e, en particular, aminoácidos, a partir dos cales se forman as proteínas, que son os compoñentes fundamentais da materia viva. É dicir, Miller conseguira formar compostos orgánicos en condicións prebiolóxicas.

O gran mérito de Miller foi, pois, demostrar que a formación de moléculas orgánicas, en condicións análogas ás da Terra primixenia, non só era posíbel, senón probábel, á vez que sentou as bases dunha nova disciplina científica, a química prebiolóxica ou química prebiótica.
En realidade, o experimento de Miller non proba de forma rotunda a teoría de Oparin, aínda que si demostra a parte máis conflitiva da mesma. En efecto, puxo de manifesto a posibilidade de que, nas condicións que supostamente se deron no noso planeta hai millóns de anos, a materia inerte puidera experimentar unha serie de transformacións que deran lugar á constitución de compostos orgánicos.
Porén, a segunda parte da teoría de Oparin, segundo a cal estas moléculas foron agrupándose durante períodos de millóns de anos até lograr adquirir as características dun ser vivo, deberán probarse mediante a teoría da evolución.
Por outra parte, o experimento de Miller serviu de punto de partida a gran número de investigadores. Seguindo a súa técnica de someter diversos compostos orgánicos a unha potente fonte de enerxía nun ambiente redutor, homes como Fox, Oró, Calvin ou Sagan, entre outros, seguiron despexando importantes incógnitas do noso pasado.
Exoxénese. A hipótese de Hoyle: a panspermia[editar | editar a fonte]
- Artigos principais: Teoría da panspermia e Fred Hoyle.

Unha alternativa á abioxénese na Terra é a hipótese de que a vida primitiva puido formarse orixinalmente fóra da Terra. A panspermia é unha hipótese que suxire que a esencia da vida está diseminada por todo o universo, e que a vida comezou na Terra grazas á chegada de tales sementes ao noso planeta.[37]
En 1879, o fisiólogo e físico alemán Hermann von Helmholtz (1821-1824), propuxo a teoría da panspermia, na que defendía que a vida tería chegado á Terra desde outro lugar do universo a través de material procedente do exterior. Este precipitaríase sobre a Terra primitiva traendo cantidades significativas de moléculas orgánicas complexas. Nos cometas hai incrustacións de capas externas que son substancias bituminosas formadas por compostos de carbono en reaccións iniciadas maiormente por luz ultravioleta.
Outra hipótese relacionada con esta é que a vida se formou en primeiro lugar no Marte primixenio, e que foi transportada á Terra cando material da súa codia foi expulsado de Marte por un asteroide e por impactos de cometas para, máis tarde, alcanzar a Terra.
A Terra estivo sometida, desde a súa formación, a un bombardeo constante de materia cósmica. Durante os primeiros 500 millóns de anos este bombardeo debeu de ser extraordinariamente intenso. As súas pegadas foron borradas pola grande actividade xeolóxica desenvolvida na súa superficie, pero na Lúa —que non tivo nin ten esta actividade— obsérvase claramente a enorme cantidade de cráteres de impacto causados polo choque de meteoritos na súa superficie. E existen estudos que suxiren a posíbel existencia de bacterias capaces de sobrevivir longos períodos de tempo incluso no espazo exterior,[38] o que apoiaría o mecanismo subxacente deste proceso.

Sobre a Terra aínda caen meteoritos, que nos permiten coñecer a composición doutros astros distintos ao noso. A análise do meteorito Murchison, así chamado por caer en 1969 na localidade dese nome, en Australia, e que foi moi ben estudado, permitiu confirmar a teoría de Hoyle —polo menos no sentido de que puideron chegar á Terra moléculas orgánicas procedentes do exterior—, pois se atopou que contiña proporcións de materia biolóxica equivalentes ás achadas no experimento de Miller, incluídas de uracilo e xantina, dous precursores das moléculas que configuran o ARN e o ADN.[39]
O maior inconveniente desta teoría é que non resolve o problema inicial de como xurdiu a vida, senón que se limita a trasladar a responsabilidade da orixe a outro lugar. Tampouco aclara como poderían ter sobrevivido as bacterias ás altísimas temperaturas e ás forzas involucradas nun impacto contra a Terra. De todos modos coñécense bacterias extremófilas capaces de soportar condicións de radiación, temperatura e presión igualmente extremas. Pese aos esforzos de Hoyle, a credibilidade da panspermia está hoxe en decadencia, el mesmo recoñeceu que a súa teoría simplemente traslada o problema a outro lugar e a outro tempo.
Os experimentos de Fox: as microesferas[editar | editar a fonte]

A publicación da teoría de Oparin e a confirmación parcial da mesma mediante o experimento de Miller deron lugar a que moitos biólogos encamiñaran as súas investigacións a descubrir como fora o longo proceso no cal a materia inerte chegou a acadar as estruturas que permitirían posteriormente o nacemento das primeira células.
Neste aspecto destacan as teorías elaboradas hai poucos anos polo bioquímico estadounidense Sidney W. Fox (1912-1998). Segundo Fox, a aparición da vida sobre o noso planeta non tivo lugar só no mar, como propoñía a teoría de Oparin, senón que tamén podería ter sucedido sobre a terra firme. Para confirmar a súa teoría fixo unha serie de experimentos, nas décadas dos 50 e os 60, nos que demostrou que, a temperaturas próximas aos 1.000 °C, unha mestura de gases similar á que supostamente formaba a atmosfera primitiva chegaba a sufrir unha serie de transformacións tales que se lograba a síntese de aminoácidos, que á súa vez se unirían entre si formando pequenos péptidos, que Fox denominou proteinoides debido ao seu parecido coas proteínas que forman parte de todos os seres vivos.
Quentou despois unha mestura destes proteinoides con auga de mar observando que, ao arrefriar, se producía un proceso de repregamento dos proteinoides sobre si mesmos, adoptando unha forma globosa, visíbel ao microscopio, polo que lle deu o nome de microesferas. Estas microesferas estarían limitadas por unha dupla capa que as protexería do exterior, e que ademais utilizarían para intercambiar materiais co medio de forma selectiva, aparecendo así o devanceiro do que posteriormente sería a membrana plasmática.
As microesferas de Fox poderían considerarse xa como entidades heterótrofas, porque a través da membrana tomarían do exterior substancias como auga, glicosa, aminoácidos etc. que, despois dunha serie de transformacións, producirían a enerxía suficiente para que continuara o desenvolvemento da microesfera. Por iso describiu este tipo de formacións como "protocélulas", capaces de crecer e de reproducirse.
Outros experimentos notábeis[editar | editar a fonte]
- Os experimentos de Joan Oró. O científico español Joan Oró obtivo nos seus experimentos sobre a orixe da vida bases nitroxenadas, que son os elementos fundamentais do ADN. En 1961 Joan Oró, engadiu ácido cianhídrico ao caldo primitivo e obtivo algunhas purinas. En 1962, noutro experimento, engadiu formaldehido e conseguiu a síntese dos azucres ribosa e desoxirribosa, compoñentes do soporte dos ácidos nucleicos no ADN e no ARN.
- Hipótese de Eigen. A principios dos anos 1970 artellouse unha grande ofensiva ao problema da orixe da vida por un equipo de científicos reunidos en torno a Manfred Eigen, do Instituto Max Planck. Intentaron examinar os estados transitorios entre o caos molecular dun caldo prebiótico e os estados transitorios dun hiperciclo de replicación, entre o caos molecular nun caldo prebiótico e sistemas macromoleculares autorreprodutores simples. Nun hiperciclo, o sistema de almacenamento de información (posibelmente ARN) produce un enzima, que cataliza a formación doutro sistema de información en secuencia até que o produto do último axuda á formación do primeiro sistema de información. Cun tratamento matemático, os hiperciclos poden crear cuasespecies que, a través da selección natural entraron nunha forma de evolución darwiniana. Un impulso á teoría do hiperciclo foi o descubrimento de que o ARN, en certas circunstancias, transfórmase en ribozimas (unha forma de enzima de ARN) capaces de catalizar as súas propias reaccións químicas.
Últimas fases da evolución prebiolóxica[editar | editar a fonte]
Como apareceu a vida?[editar | editar a fonte]
- Artigos principais: Bases nitroxenadas, Ácido ribonucleico e Ácido desoxirribonucleico.

Xa foran as microesferas de Fox, os coacervados de Oparin na auga, ambas as entidades ao mesmo tempo, ou outras formas non determinadas, nun momento dado aparecerían sobre o noso planeta unha serie de estruturas que tiñan un grao de complexidade moito maior có resto da materia que as arrodeaba.
Unha vez aparecidas comezou a actuar sobre elas o mecanismo da evolución e a selección natural, mediante a cal moitas desaparecerían mentres que outras, as que máis desenvolvida tiñan a súa estrutura, continuarían medrando e facéndose cada vez máis complexas.
Non obstante, é probábel que estes cambios non tivesen lugar dunha forma lenta e paulatina, senón mediante bruscas variacións, producíndose no interior das mesmas novas e máis complexas substancias orgánicas, e pequenos orgánulos capaces de realizar funcións cada vez máis difíciles.
E, tras unha última transformación, converteríanse en células capaces de desenvolver xa de forma definitiva todas as funcións vitais.
Seguramente os procesos da evolución prebiolóxica nunca poderán ser demostrados, e a súa existencia quedará reducida a unha teoría con maior ou menor número de probas ao seu favor ou na súa contra, xa que un proceso coma este será practicamente imposíbel de reproducir experimentalmente. Non obstante, a evolución prebiolóxica conta coa suficiente cantidade de probas ao seu favor como para que na actualidade sexa considerada válida pola maior parte da comunidade científica. Aínda que non teñamos datos inequívocos de que sucedeu hai tanto tempo, podemos tratar de reconstruír o pasado grazas a indicios sobre o comportamento das moléculas en condicións simuladas semellantes ás que supostamente reinaban nos inicios da Terra. De querermos analizar como apareceu a vida na Terra, debemos partir dunha definición de ser vivo como a seguinte:
Un ser vivo é un ser capaz de de autorreplicarse e de evolucionar co tempo mediante a selección natural.
E se buscamos un sistema capaz de cumprir estas características comprobaremos que, de todas as moléculas posíbeis, a mellor candidata a molécula inicial é o ácido ribonucleico (ARN). A hipótese máis aceptada actualmente é a seguinte:
O caldo primitivo íase arrequecendo a medida que caían os compostos producidos na atmosfera. De todos estes, os nucleótidos tiñan a propiedade de unírense entre si formando compostos estábeis. O azar puido determinar que algúns destes agrupamentos se uniran á vez para formar moléculas máis grandes, os polinucleótidos. Posibelmente así construiríase a primeira molécula de ARN. Certas moléculas de ARN adquirirían a capacidade de dirixir a síntese de proteínas. Por outra parte, algunhas destas moléculas acadarían a propiedade de facilitar a formación de copias idénticas de si mesmas (de replicarse).
O ARN está hoxe considerado como mellor candidato a primeira molécula biolóxica porque ten información para facer copias de si mesmo e os científicos pensan que a propia molécula podería dirixir o proceso, aínda que se descoñece como puido formarse dito ARN. Este ácido nucleico está composto por catro tipos de ribonucleótidos diferentes nos que o único que cambia duns a outros é a base nitroxenada, que pode ser adenina (A), guanina (G), citosina (C) ou uracilo (U). Os outros compoñentes son ribosa e ácido fosfórico.
-
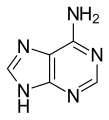 Adenina
Adenina -
 Guanina
Guanina -
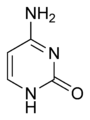 Citosina
Citosina -
 Uracilo
Uracilo -
 Timina
Timina





Estrutura química das bases nitroxenadas. No ADN a timina substitúe ao uracilo do ARN.
As moléculas de ARN constrúense todas do mesmo modo, pero pode cambiar a orde na que se coloquen as bases, orixinando así diferentes mensaxes biolóxicas. Por outra parte, demostrouse recentemente que moléculas de ARN poden catalizar certas reaccións (actividade de ribozima). Isto é o que fai que moitos biólogos pensen que o ARN foi a primeira molécula capaz de autorreplicarse. Pero, no proceso de copiarse a si mesma ou autorreplicación, prodúcense con frecuencia cambios ou erros na orde de colocación das bases nitroxenadas. Estes erros, que son herdados polas moléculas fillas, chamámolos mutacións. As mutacións son a fonte da variabilidade dos seres vivos.
A maioría destes erros producirían moléculas aparentemente normais no seu funcionamento, ou quizais defectuosas, pero algunhas destas mutacións, por azar, puido dar lugar a moléculas con mellor capacidade de reprodución. Entre todas as variantes, aquelas que se reproduciran con máis eficiencia darían lugar a máis copias de si mesmas, desprazando lentamente ás menos eficientes: aparecería deste xeito a selección natural como mecanismo evolutivo para escoller as variantes mellor adaptadas. Debido á presenza da membrana externa, as proteínas sintetizadas por unha determinada molécula de ARN quedaban retidas e dispoñíbeis só para o ARN que as orixinara, o que aumentaría a súa eficacia. Estas vesículas, que nun momento determinado puideron estrangularse e dividirse, deberon ser as precursoras das primeiras células.
Estas células primitivas deberon de ser moi pequenas, e de forma esférica. Conterían enzimas e proteínas, e a súa información xenética localizaríase no ARN. Talvez consumían moitos tipos de moléculas orgánicas, como glícidos, nucleótidos e aminoácidos. Nalgún momento do proceso evolutivo produciríase a aparición do ADN ou ácido desoxirribonucleico, que substituíu o ARN, na maior parte dos organismos, na función de almacenar a información para a síntese de proteínas.
Evolución biolóxica: cara á célula eucariota[editar | editar a fonte]
- Véxase tamén: Estromatólito, Evolución das células e Eucarioxénese.

Hoxe sabemos que a vida apareceu na Terra hai uns 4.000 millóns de anos. En Warrawoona (Australia)[40] hai xa anos J. W. Shopt atopou unhas formacións rochosas en forma de columnas ou de cúpulas chamadas estromatólitos que tiñan unha antigüidade duns 3.500 millóns de anos e, canda elas, fósiles que recordan fileiras de células semellantes ás das algas azuis ou cianobacterias.
Por outra parte, en Groenlandia atopáronse rochas sedimentarias formadas hai máis de 3.800 millóns de anos (son as máis antigas das encontradas até agora). A presenza de rochas sedimentarias proba que naquela época xa existía auga, indispensábel para a vida. Ademais, nestas rochas acháronse indicios da presenza de carbono.
De calquera forma, parece que a vida puido orixinarse hai uns 3.800 millóns de anos con células semellantes ás das bacterias actuais.
A evolución posterior destas células primitivas pode esquematizarse do seguinte xeito:
- As primeiras células, procariotas, nutriríanse de materia orgánica no caldo primitivo, descompoñendo, por exemplo, azucres mediante fermentacións, sendo, polo tanto, heterótrofas anaerobias, xa que aínda non había oxíxeno na atmosfera terrestre.
- A medida que a materia orgánica foi escaseando, a evolución por selección natural conduciría á aparición da capacidade para fixar nitróxeno atmosférico. Estas primitivas células terían tamén mecanismos de reparación do seu material xenético (ADN), xa que os raios ultravioleta —que causan mutacións (cambios) no ADN— chegarían con grande intensidade á superficie da Terra ao non existir aínda o filtro da capa de ozono.
- Algunhas destas células desenvolverían a capacidade de producir pigmentos captadores de enerxía luminosa. Comezaría así a fotosíntese, pero sen produción de osíxeno, xa que a fonte de hidróxeno serían compostos de xofre, ou mesmo H2 atmosférico, non a auga. Trataríase, por tanto, de organismos fotosintetizadores anaerobios.
- O seguinte paso daríano aquelas células que puideran obter o hidróxeno mediante a fotólise da auga. Deste xeito aparecería a fotosíntese con produción de osíxeno. Este feito sería transcendental para a evolución posterior da vida, xa que pouco a pouco foi diminuíndo a cantidade de dióxido de carbono (CO2) na atmosfera e aumentando a cantidade de osíxeno (O2: a atmosfera puido así pasar de ser redutora a oxidante, abrindo a posibilidade para a existencia de seres aerobios. Estes organismos serían as cianobacterias.
- A gran competencia que sufrirían as bacterias anaerobias faría que algunhas destas evolucionaran cara á adquisición dun metabolismo aerobio, posibilitado pola presenza de osíxeno na atmosfera. Nacería así a respiración, coas primeiras bacterias aerobias.
- Por último, hai uns 1.800 millóns de anos, xurdiron as primeiras células eucariotas, dotadas de compartimentos con membrana, como o núcleo, que encerra o material xenético. A súa complexidade e alto grao de organización supuxo outro cambio importante na evolución.
- Algúns efectos da "revolución do osíxeno"
- 1. A atmosfera pasou de ser redutora a ser oxidante.
- 2. O metano, probabelmente, reaccionou co osíxeno dando dióxido de carbono e auga:
- CH4 + O2 → 2 H2O
- 3. O amoníaco, en presenza de osíxeno, pasou a formar nitróxeno e auga:
- 4 NH3 + 3 O2 → 2 N2 + 6 H2O
- 4. Ao combinárense entre si as moléculas de osíxeno, baixo as radiacións de alta enerxía, formaríase ozono:
- O2 + 2 O2 → 2 O3
- 5. Os metais puros e outras substancias minerais da codia terrestre, ao reaccionaren co osíxeno, convertéronse en óxidos, constituíntes dos minerais e rochas que se achan na superficie do noso planeta.
Teoría endosimbiótica[editar | editar a fonte]


- Artigos principais: Teoría endosimbiótica e Lynn Margulis.
As causas e a maneira de producirse esta evolución cara á célula eucariota explícanse hoxe por dúas teorías principais: a autóxena e a endosimbiótica.
Segundo a teoría autóxena, formulada por F. J. A. Taylor e E. D. Dodson, o incremento progresivo do tamaño das células iría acompañado dun desenvolvemento de compartimentos no citoplasma, para o que a membrana celular se invaxinaría e iría formando a membrana nuclear e os demais compartimentos membranosos especializados (retículo endoplasmático, aparello de Golgi, mitocondrias, cloroplastos). Aínda que ten ao se favor, entre outras cousas, o feito de que todas as membranas celulares teñen a mesma estrutura —o que suxire unha orixe común, a partir da plasmática— ao non observárense na actualidade formas intermedias deste proceso, esta teoría non recibe hoxe moitas adhesións.
De acordo coa teoría endosimbiótica, debida á bióloga estadounidense Lynn Margulis (nada en 1938), unha gran bacteria anaerobia fagocitaría e incorporaría, sen dixerila, outra bacteria aerobia, pero de menor tamaño, establecéndose entre elas unha relación simbiótica, de maneira que a bacteria pequena acaba transformándose nunha mitocondria. Así podería formarse a primeira célula eucariótica heterótrofa, que podería dar lugar ás células dos animais e dos fungos.
Outro proceso de endosimbiose puido establecerse entre a célula eucariótica así formada e outra do tipo das cianobacterias actuais (autótrofa). Esta, máis pequena, podería converterse nun cloroplasto, formándose así unha célula antecesora das algas e dos vexetais. Posteriormente incorporaríanse bacterias do tipo das espiroquetas, que se converterían en centríolos, flaxelos e fibrilas. Estas novas estruturas intervirían na ordenación dos elementos do citoplasma. E o centríolo, ao longo do tempo, chegaría a controlar a ordenación dos cromosomas, o que podería ser a orixe da mitose.
A teoría da endosimbiose, ou da endosimbiose seriada, como tamén se a coñece, é a que conta con maior apoio da comunidade científica na actualidade. As principais probas ao seu favor son:
- As mitocondrias e os cloroplastos posúen o seu propio material xenético, formado por un cromosoma circular, e os seus propios ribosomas, semellantes aos ribosomas das células eucarióticas.
- As mitocondrias e os cloroplastos son capaces de realizar a síntese de proteínas a escala limitada.
- As mitocondrias e os cloroplastos poden destruírse con antibióticos que matan bacterias, pero non células eucarióticas.
- Na actualidade dánse algunhas relacións endosimbióticas. No intestino dos termes vive un protozoo, Myxotrichia paradoxa, que á súa vez ten outros endosimbiontes, como bacterias, que actúan de flaxelos. A maioría de células coralinas teñen no seu interior algas, o que fai posíbel que estes arrecifes sexan tan produtivos.
En todo caso, a teoría endosimbiótica non resolve completamente todos os interrogantes de como xurdiron as células eucarióticas a partir das procarióticas. Tampouco non explica como o material xenético do núcleo chegou a rodearse dunha membrana.
Modelos actuais[editar | editar a fonte]
Non existe un modelo xeneralizado da orixe da vida. Os modelos actualmente máis aceptados constrúense dunha ou doutra maneira sobre certo número de descubrimentos acerca da orixe dos compoñentes moleculares e celulares da vida, enumerados na orde máis ou menos aproximada na que se postularon:
- As posíbeis condicións prebióticas remataron coa creación de certas moléculas pequenas básicas (monómeros) da vida, como os aminoácidos. Isto demostrouse no experimento de Miller, en 1953.
- Os fosfolípidos (dunha lonxitude adecuada) poden formar espontaneamente bicapas lipídicas, un dos dous compoñentes básicos da membrana celular.
- A polimerización dos nucleótidos en moléculas de ARN ao azar puido dar lugar a ribozimas autorreplicantes (hipótese do mundo de ARN).
- As presións de selección para unha eficiencia catalítica e unha diversidade maior darían lugar a ribozimas que catalizaban a transferencia de péptidos (e, por ende, a formación de pequenas proteínas), xa que os oligopéptidos formarían complexos co ARN para formar mellores catalizadores. Deste xeito xurdiría o primeiro ribosoma e a síntese de proteínas faríase máis prevalente.
- As proteínas superan aos ribozimas na súa capacidade catalítica e por tanto convértense no biopolímero dominante. Os ácidos nucleicos quedan restrinxidos a un uso predominantemente xenómico.
A orixe das biomoléculas básicas, aínda que non se estableceu completamente, é menos controvertido que o significado e orde dos pasos 2 e 3. Os reactivos químicos inorgánicos básicos a partir dos cales se formou a vida son metano, amoníaco, auga, sulfuro de hidróxeno (H2S), dióxido de carbono e anión fosfato.
Aínda non se sintetizou unha protocélula utilizando os compoñentes básicos que teña as propiedades necesarias para a vida (o chamado enfoque "de abaixo a arriba"). Sen esta proba de principio, as explicacións tenden a quedar curtas. Non obstante, algúns investigadores están traballando neste campo como, por exemplo, Jack Szostak da Universidade Harvard. Outros autores argumentaron que un enfoque "de arriba a abaixo" sería máis probábel. Un destes intentos realizouno Craig Venter e colaboradores no Institute for Genomic Research. Utilizaba enxeñería xenética con células procariotas cunha cantidade de xenes progresivamente menor, intentando discernir en que punto se alcanzaban os requisitos mínimos para a vida. O biólogo John Desmon Bernal cuñou o termo biopoese para este proceso, e suxiriu que había un número de "estadios" claramente definidos que se podían recoñecer á hora de explicar a orixe da vida:
- Estadio 1: A orixe dos monómeros biolóxicos.
- Estadio 2: A orixe dos polímeros biolóxicos.
- Estadio 3: A evolución desde o molecular cara á célula.
Bernal suxeriu que a evolución darwiniana puido comezar moi cedo, nalgún momento entre os dous primeiros estadios listados. Pódese argumentar que o cambio máis crucial que aínda segue sen recibir resposta por esta teoría é como estes "ladrillos" orgánicos relativamente simples polimerizan forman estruturas máis complexas, interactuando de modo consistente para formar unha protocélula. Por exemplo, nun ambiente acuoso, a hidrólise de oligómeros/polímeros nos seus constituíntes monoméricos está enerxeticamente favorecida sobre a condensación de monómeros individuais en polímeros. Ademais, o experimento de Miller produce moitas substancias que acabarían dando reaccións cruzadas cos aminoácidos ou terminando a cadea peptídica.
Hipótese de Wächstershäuser[editar | editar a fonte]
- Artigo principal: Teoría do mundo de ferro-xofre.

Outra posíbel resposta a este misterio da polimerización propúxoa Günter Wächtershäuser en 1980, na súa teoría do ferro-xofre.[43] Nela postulou a evolución das rotas (bio)químicas como o fundamento da evolución da vida. Incluso presentou un sistema consistente para rastrexar as pegadas da actual bioquímica desde as reaccións ancestrais que proporcionaban rotas alternativas para a síntese de "ladrillos orgánicos" a partir de compoñentes gasosos simples.
Ao contrario que nos experimentos clásicos de Miller, que dependían de fontes externas de enerxía (como relampos simulados ou irradiación UV), os "sistemas de Wächstershäuser" veñen cunha fonte de enerxía incorporada, os sulfuros de ferro e outros minerais (por exemplo a pirita). A enerxía liberada a partir das reaccións redox deses sulfuros metálicos non só estaría dispoñíbel para a síntese de moléculas orgánicas, senón tamén para a formación de oligómeros e polímeros. Propón por iso a hipótese de que tales sistemas poderían ser capaces de evolucionar até formaren conxuntos autocatalíticos de entidades autorreplicantes metabolicamente activas, que serían as precursoras das actuais formas de vida.
O experimento tal e como foi levado a cabo rendeu unha produción relativamente pequena de dipéptidos (do 0,4 % ao 12,5 %) e unha produción aínda moi inferior de tripéptidos (0,003 %), e os autores advirtiron de que "baixo estas mesmas condicións os dipéptidos se hidrolizaban rapidamente"[44]. Outra crítica do resultado é que o experimento non incluía ningunha organomolécula que puidera con maior probabilidade dar reaccións cruzadas ou terminar a cadea (Huber e Wächsterhäuser, 1998).
A última modificación da hipótese do ferro-xofre propuxérona William Martin e Michael Russell en 2002.[45] De acordo co seu escenario, as primeiras formas celulares de vida puideron evolucionar dentro das chamadas "fontes hidrotermais" (chemineas ou fumarolas negras) nas profundidades onde se encontran as zonas de expansión do fondo oceánico. Estas estruturas consisten en diminutas cavernas que están revestidas por delgadas paredes membranosas de sulfuros metálicos. Por tanto, estas estruturas resolverían varios puntos críticos dos sistemas de Wächstershäuser "puros" dunha soa vez:
- As microcavernas proporcionan medios para concentrar as moléculas acabadas de sintetizar, por tanto aumentando a posibilidade de formar oligómeros.
- Os abruptos gradientes de temperatura existentes dentro dunha cheminea negra permiten establecer "zonas óptimas" de reaccións parciais en diferentes rexións da mesma (por exemplo, a síntese de monómeros nas zonas máis quentes e a oligomerización nas zonas máis frías).
- O fluxo de auga hidrotermal a través da estrutura subministra unha fonte constante de "ladrillos" e enerxía (sulfuros metálicos acabados de precipitar).
- O modelo permite unha sucesión de diferentes pasos de evolución celular (química prebiótica, sínteses de monómeros e oligómeros, sínteses de péptidos e proteínas, mundo de ARN, ensamblaxe de ribonucleoproteínas e mundo de ADN) nunha única estrutura, facilitando o intercambio entre todos os estadios de desenvolvemento.
- A síntese de lípidos como medio de "illar" as células do medio ambiente non é necesaria até que estean todas as funcións celulares basicamente desenvolvidas.
Este modelo sitúa ao último antepasado común universal (LUCA, do inglés Last Universal Common Ancestor) dentro dunha cheminea negra, en lugar de asumir a existencia dunha forma de vida libre de LUCA. O último paso evolutivo sería a síntese dunha membrana lipídica que finalmente permitira ao organismo abandonar o sistema no interior da microcaverna das fumarolas negras e comezar a súa vida independente. Este postulado dunha adquisición tardía dos lípidos é consistente coa presenza de tipos completamente diferentes de lípidos de membrana en arqueobacterias e eubacterias (máis os eucariotas) cunha fisioloxía moi similar en todas as formas de vida noutros aspectos.
Outro asunto sen resolver na evolución química é a orixe da homoquiralidade. Por exemplo, todos os monómeros teñen a mesma "man dominante" (os aminoácidos son zurdos e os ácidos nucleicos e azucres son destros). A homoquiralidade é esencial para a formación de ribozimas funcionais (e probabelmente tamén de proteínas). A orixe da homoquiralidade podería explicarse simplemente por unha asimetría inicial por casualidade seguida dunha descendencia común.
Os traballos levados a cabo en 2003 por científicos de Purdue identificaron o aminoácido serina como a probábel raíz que provoca a homoquiralidade das moléculas. A serina forma enlaces particularmente fortes cos aminoácidos da mesma quiralidade, o cal dá lugar a un grupo de oito moléculas que poderían todas elas ser destras ou zurdas. Esta propiedade contraponse á doutros aminoácidos que son capaces de formar enlaces débiles cos aminoácidos de quiralidade oposta. Aínda que o misterio de por que acabou sendo dominante a serina zurda está sen resolver, os resultados suxiren unha resposta á cuestión da transmisión quiral: o como as moléculas orgánicas dunha quiralidade manteñen a dominancia unha vez que se establece a asimetría.
Teoría da praia radioactiva[editar | editar a fonte]
Zachary Adam,[46] da Universidade de Washington en Seattle, afirma que procesos mareais maiores que os actuais, producidos por unha Lúa situada a unha distancia moito menor, poderían concentrar partículas radioactivas de uranio e outros elementos radioactivos na marea alta nas praias primordiais onde deberon ser os responsábeis de xerar os compoñentes elementais da vida. De acordo cos modelos de computación publicados en Astrobiology[47] un depósito de tales materiais radioactivos podería ter a mesma reacción nuclear autosostida que se dá no xacemento de uranio de Oklo, en Gabón. Esta area radioactiva subministra suficiente enerxía como para xerar moléculas orgánicas, como aminoácidos e azucres a partir de acetonitrilo procedente da auga. A monazita radioactiva tamén libera fosfatos solúbeis nos espazos que se encontran entre os grans de area, facéndoos bioloxicamente accesíbeis. Así pois os aminoácidos, azucres e fosfatos solúbeis poden producirse simultaneamente, de acordo con Adam. Os actínidos radioactivos, que entón se encontraban en maiores concentracións, puideron formar parte de complexos órganometálicos. Estes complexos puideron ser importantes como primeiros catalizadores nos procesos da vida.
John Parnell, da Universidade de Aberdeen, suxire que tales procesos formaron parte do "crisol da vida" nos comezos de calquera planeta rochoso até que este fora o suficientemente grande como para xerar un sistema de tectónica de placas que achegara minerais radioactivos á superficie. Posto que se cre que a Terra na súa orixe estaba formada por moitas "microplacas", daríanse condicións favorábeis para este tipo de procesos.
Homoquiralidade[editar | editar a fonte]
Algúns procesos da evolución química deberían explicar a orixe da homoquiralidade, é dicir, o feito de que todos os compoñentes elementais dos seres vivos teñen a mesma quiralidade, sendo os aminoácidos levoxiros, e os azucres ribosa e desoxirribosa dos ácidos nucleicos dextroxiros, así como os fosfoglicéridos quirais. Pódense sintetizar moléculas quirais, pero en ausencia dunha fonte de quiralidade ou dun catalizador quiral, fórmase unha mestura 50/50 de ambos os enantiómeros, á cal se lle chama mestura racémica. Clark suxeriu que a homoquiralidade puido comezar no espazo, posto que os estudos sobre os aminoácidos do meteorito Murchison amosaron que a L-alanina era dúas veces máis frecuente que a forma D, e o ácido L-glutámico era tres veces máis prevalente que a súa contrapartida dextroxira. Suxeriuse que a luz polarizada tivo o poder de destruír un dos enantiómeros dentro do disco protoplanetario. Noyes demostrou que a desintegración beta provocaba a destrución da D-leucina nunha mestura racémica, e que a presenza de 14C, presente en grandes cantidades nas substancias orgánicas do ambiente primitivo da Terra, podería ser a causa.[48] Robert M. Hazen publicou informes de experimentos realizados en distintas superficies cristalinas quirais que actuaban como posíbeis lugares de concentración e ensamblamento de monómeros quirais en macromoléculas.[49] Unha vez estabilizado o sistema, a quiralidade podería seleccionarse favorabelmente pola evolución.[50] Os traballos con compostos orgánicos encontrados en meteoritos tenden a suxerir que a quiralidade é unha característica da síntese abioxénica, posto que os aminoácidos actuais son levoxiros, mentres que os azucres son predominantemente dextroxiros.[51]
Autoorganización e replicación[editar | editar a fonte]
- Artigo principal: Autoorganización.
Se se considera, como a miúdo sucede, que a autoorganización e a autorreplicación son os procesos principais que caracterizan aos sistemas vivos, cómpre dicir que hai moitos tipos de moléculas abióticas que exhiben estas características nas condicións adecuadas. Por exemplo, Martin e Russel demostraron que a formación dun compartimento distinto do ambiente por membranas celulares e a autoorganización de reaccións redox autocontidas son os atributos máis conservados entre os seres vivos, e isto lévaos a argumentar que a materia inorgánica con estes atributos poderían estar entre os atributos máis probábeis do antepasado común de todos os seres vivos.[45]
Das moléculas orgánicas ás protocélulas[editar | editar a fonte]
A cuestión de como unhas moléculas orgánicas simples forman unha protocélula leva moito tempo sen resposta, pero existen moitas hipóteses. Algunhas destas postulan unha temperá aparición dos ácidos nucleicos (teorías denominadas "primeiro os xenes") mentres que outras postulan que primeiro apareceron as reaccións bioquímicas e as rotas metabólicas (as "primeiro o metabolismo"). Recentemente apareceron tendencias con modelos híbridos que combinan aspectos de ambas as dúas.
Modelos "primeiro os xenes": o mundo de ARN[editar | editar a fonte]
- Artigo principal: Hipótese do mundo de ARN.

A hipótese do mundo de ARN enunciouna Walter Gilbert, da Universidade Harvard, con base nos experimentos de Thomas Cech (Universidade de Colorado) e Sidney Altman (Universidade Yale) en 1980. Suxiren que as moléculas relativamente curtas de ARN poderían formarse espontaneamente de xeito que foran capaces de catalizar a súa propia replicación continua.[52] É difícil calibrar a probabilidade desta formación. Expuxéronse algunhas hipóteses de como puido suceder isto. As primeiras membranas celulares puideron formarse espontaneamente a partir de proteinoides (moléculas similares a proteínas que se producen cando se quentan solucións de aminoácidos). Cando están á concentración correcta en solución acuosa forman microesferas que, segundo se observou, presentan unha conduta similar á dos compartimentos rodeados de membrana.
Outras posibilidades inclúen sistemas de reaccións químicas que teñen lugar no interior de susbtratos de arxila ou na superficie de rochas piríticas. Entre os factores que apoian un papel importante para o ARN na vida primitiva inclúese a súa habilidade para replicarse; a súa habilidade tanto para almacenar información coma para catalizar reaccións químicas (como ribozimas); o seu papel importantísimo como intermediario na expresión e mantemento da información xenética (en forma de ADN) nos organismos modernos e na facilidade da súa síntese química ou, polo menos, dos compoñentes da molécula baixo as condicións aproximadas da Terra primitiva. Producíronse artificialmente no laboratorio moléculas de ARN relativamente curtas capaces de duplicar a outras.[53]
Un punto de vista lixeiramente distinto sobre esta mesma hipótese é a de que un tipo diferente de ácido nucleico, como os ácidos nucleicos peptídicos (ANP) ou os ácidos nucleicos de treosa (TNA) foron os primeiros en emerxer como moléculas autorreprodutoras para seren despois substituídas polo ARN.[54][55]
Aínda quedan algúns problemas coa hipótese do mundo de ARN, en particular a inestabilidade do ARN cando se expón á radiación ultravioleta, a dificultade de activar e ligar os nucleótidos, a carencia de fosfato dispoñíbel en solución, requirida para construír a súa columna vertebral, e a inestabilidade da base citosina (que é susceptíbel á hidrólise). Recentes experimentos tamén suxiren que as estimacións orixinais do tamaño dunha molécula de ARN capaz de autorreplicación eran moi probabelmente amplamente subestimadas. Formas máis modernas da teoría do mundo de ARN propoñen que unha simple molécula era capaz de autorreplicación (que outro "mundo" por tanto evolucionou co tempo até producir o mundo de ARN). Neste momento, non obstante, as distintas hipóteses teñen insuficientes probas que o apoien. Moitas destas poden ser simuladas e probadas no laboratorio, pero a ausencia de rochas sedimentarias sen alterar desde un momento tan inicial na historia da Terra deixa poucas oportunidades de probar adecuadamente esta hipótese.
Modelos "primeiro o metabolismo": o mundo de ferro-xofre e outros[editar | editar a fonte]
- Artigo principal: Teoría do mundo de ferro-xofre.

Algúns modelos rexeitan a idea da autorreplicación dun "xene espido" e postulan a aparición dun metabolismo primitivo que puido proporcionar un ambiente para a posterior emerxencia da replicación do ARN. Unha das primeiras encarnacións desta idea foi a presentada en 1924 pola noción de Alexander Oparin de primitivas vesículas autorreplicantes que precederon ao descubrimento da estrutura do ADN. As variantes máis recentes dos anos 1980 e 1990 inclúen a teoría do mundo de ferro-xofre de Günter Wächtershäuser e modelos presentados por Christian de Duve baseados na química dos tioésteres.
Entre algúns modelos máis abstractos e teóricos da plausibilidade da emerxencia do metabolismo sen a presenza de xenes inclúese un modelo matemático presentado por Freeman Dyson a principios dos anos 1980 e a noción de Stuart Kauffman de conxuntos colectivamente autocatalíticos, discutidos xa avanzada a década. Porén, a idea de que un ciclo metabólico pechado, como o ciclo redutor do ácido cítrico proposto por Günter Wächstershäuser, puido formarse espontaneamente, aínda permanece sen probas. De acordo con Leslie Orgel, un líder nos estudos sobre a orixe da vida durante algunhas das pasadas décadas, hai razóns para crer que a afirmación permanecerá así. Nun artigo titulado Self-Organizing Biochemichal Cycles Orgel resume a súa análise da proposta estipulando que "polo de agora non existe razón para esperar que ciclos de múltiples pasos como o ciclo redutor do ácido cítrico puidera autoorganizarse na superficie de FeS/FeS2 ou dalgún outro mineral".[57] É posíbel que outro tipo de rota metabólica se usara nos comezos da vida. Por exemplo, en lugar do ciclo redutor do ácido cítrico, a rota aberta do acetil-CoA (outra das catro vías recoñecidas de fixación de dióxido de carbono na natureza actualmente) podería ser máis compatíbel coa idea de autoorganización nunha superficie de sulfuro metálico. O enzima clave desta vía, monóxido de carbono deshidroxenase/acetil-CoA, ten ancorados grupos mixtos de sulfuro de ferro e níquel nos seus centros de reacción e cataliza a formación de acetil-CoA (que podería considerarse como unha forma moderna de acetilo-tiol) nun único paso.
Teoría da burbulla[editar | editar a fonte]
As ondas que rompen nas costas crean unha delicada escuma composta por burbullas. Os ventos que varren o océano teñen tendencia a levar cousas á costa, de forma similar á madeira que se amorea á deriva nunha praia. É posíbel que as moléculas orgánicas puideran concentrarse nos bordos costeiros dun xeito parecido. As augas costeiras máis superficiais tamén tenden a seren máis cálidas, concentrando máis tarde as moléculas orgánicas por evaporación. Mentres as burbullas formadas maiormente por auga estalan rapidamente, ocorre que as burbullas de graxa son moito máis estábeis, dándolle máis tempo a cada burbulla en particular para levar a cabo estes cruciais experimentos.
Os fosfolípidos son un bo exemplo dun composto graxo que se cre que foi prevalente nos mares prebióticos. Debido a que os fosfolípidos teñen unha cabeza hidrofílica nun extremo e unha cola hidrofóbica no outro, teñen tendencia a formar espontaneamente bicapas lipídicas en auga. Unha burbulla de monocapa lipídica só pode conter graxa, mentres que unha burbulla de bicapa lipídica só pode conter auga, sendo un probábel precursor das modernas membranas celulares. Se unha proteína acaba incrementando a integridade da súa burbulla nutriz, entón a burbulla ten unha gran vantaxe e acaba situándose na cúspide da selección natural. A primitiva reprodución podería visualizarse cando as burbullas estalaban, liberando o resultado do experimento no seu medio circundante. Unha vez que se libera unha cantidade suficiente do "material correcto", poderíase lograr o desenvolvemento dos primeiros procariotas, eucariotas e organismos multicelulares.[58] De modo similar, as burbullas formadas unicamente por moléculas similares a proteínas, chamadas microesferas, formaríanse espontaneamente baixo as condicións adecuadas. Pero non hai precursores probábeis das modernas membranas celulares, posto que as membranas celulares están compostas primariamente de compoñentes lipídicos máis que de compoñentes aminoacídicos.
Un modelo recente posto a punto por Fernando e Rowe suxire que o confinamento dun metabolismo autocatalítico non-enzimático dentro das protocélulas podería ser un modo de evitar o problema das reaccións colaterais que son típicas dos modelos de "metabolismo primeiro".
Modelos híbridos[editar | editar a fonte]
Unha crecente comprensión dos erros dos modelos que consideran puramente "primeiro os xenes" ou "primeiro o metabolismo" está levando a tendencias cara a modelos que incorporan aspectos de ambos. Concretamente e, baseándose na implausibilidade loxística do modelo do mundo de ARN, Robert Griffith estableceu un modelo de cooperación entre o ARN, os primitivos péptidos e lípidos cuxa secuencia sería como segue: a materia orgánica xerada mediante síntese abiótica nos océanos primitivos separaríase en fases por densidade e solubilidade. A síntese de monómeros complexos e tamén de polímeros tería lugar en superficies hidrofílicas e nas súas proximidades (interfases) tanto no medio acuoso coma no aéreo. A replicación e a tradución encontrarían o seu lugar idóneo na interfase de elementos emulsificados de carácter hidrofóbico. Os primeiros ácidos nucleicos codificarían preferentemente arxinina. Estes péptidos arrequecidos en arxinina servirían para secuestrar e transferir enlaces de fosfato ricos en enerxía. Isto faría que os péptidos foran esenciais para o mantemento dos ácidos nucleicos, ricos en fosfato, e ao mesmo tempo manteríanos cerca das interfases lipídicas.[59]
Outros modelos[editar | editar a fonte]
Autocatálise[editar | editar a fonte]
O etólogo británico Richard Dawkins escribiu sobre a autocatálise como unha explicación potencial para a orixe da vida no seu libro A historia do antepasado (2004). Os autocatalizadores son substancias que catalizan a súa propia produción e teñen por tanto a propiedade de ser un replicador molecular simple. Neste libro, Dawkins cita experimentos levados a cabo por Julius Rebek e colaboradores no Sripps Research Institute de California no que combinan aminoadenosina e éster de pentafluorofenilo co autocatalizador éster triácido de aminoadenosina (AATE). Un sistema do experimento contiña variantes de AATE que catalizaban a súa propia síntese. Este experimento demostraba a posibilidade de que a autocatálise podía ser competente nunha poboación de entidades con herdanza, que podería interpretarse como unha forma rudimentaria de selección natural.
Teoría da arxila[editar | editar a fonte]
Graham Cairns-Smith, da Universidade de Glasgow, presentara unha hipótese sobre a orixe da vida en 1985 baseada na arxila que foi adoptada como unha explicación plausíbel por só uns poucos científicos (incluíndo a Richard Dawkins). A teoría da arxila postula que as moléculas orgánicas complexas creceron gradualmente nunha plataforma de replicación non orgánica preexistente —cristais de silicato en disolución—. A complexidade das moléculas acompañantes que se desenvolvía como unha función das presións de selección en tipos de cristais de arxila extráese entón para servir á replicación de moléculas orgánicas independentemente da súa "pista de engalaxe" no seu silicato.
Cairns-Smith é un firme crítico doutros modelos de evolución química.[60] Non obstante admite que, como moitos modelos da orixe da vida, o seu tamén ten defectos (Horgan 1991). É verdadeiramente "sacar a vida de debaixo das pedras".
Peggy Rigou do Instituto Nacional de Investigación Agronómica de EE. UU. (INRA), en Jouy-en-Josas, Francia, publicou na edición do 11 de febreiro de Science News que os prións son capaces de unirse a partículas de arxila e abandonar estas partículas cando a arxila se carga negativamente. Aínda que non fai ningunha referencia ás implicacións para as teorías da orixe da vida, esta investigación podería suxerir que os prións son unha rota probábel cara ás primeiras moléculas reprodutoras. En 2007, Kahr e colaboradores publicaron os seus experimentos que examinan a idea de que os cristais poden actuar como unha fonte de información transferíbel, usando cristais de ftalato de potasio hidroxenado. Os cristais "nai" con imperfeccións cortáronse e usáronse como sementes para criar cristais "fillos" a partir da solución. Entón examinaron a distribución das imperfeccións no sistema cristalino e encontraron que as imperfeccións dos cristais nai realmente reproducíanse nos fillos. Os cristais fillos tiñan moitas imperfeccións adicionais. Para unha conduta paraxenética as imperfeccións adicionais deberían ser moito menores que as dos pais, e de aí que Kahr conclúa que os cristais "non eran o suficientemente fieis como para almacenar información dunha xeración á seguinte".[61][62]
Modelo de Gold de "Biosfera profunda e quente"[editar | editar a fonte]
O descubrimento dos nanobios (estruturas filamentosas máis pequenas que as bacterias que conteñen ADN) en rochas profundas, levou a unha teoría controvertida presentada por Thomas Gold a principios dos anos 1990 na que se expoñía que a vida se desenvolveu ao principio non na superficie da Terra, senón varios quilómetros baixo a superficie. Agora sábese que a vida microbiana é abundante a máis de cinco quilómetros baixo a superficie da Terra en forma de arqueobacterias, que se considera que se orixinaron ou antes ou aproximadamente ao mesmo tempo que as eubacterias, moitas das cales viven na superficie (incluíndo os océanos). Afirmou que o descubrimento de vida microbiana baixo a superficie doutro corpo do sistema solar daría un creto significativo a esta teoría. Tamén dicía que unha subministración de nutrientes dunha fonte profunda e inalcanzábel promovería a supervivencia porque a vida que xorde nun amoreamento de materia orgánica probabelmente consumiría todo o seu alimento e acabaría extinguíndose.
O mundo de lípidos[editar | editar a fonte]
Hai unha teoría que afirma que as primeiras substancias autorreplicantes eran de tipo lipídico.[63] Sábese que os fosfolípidos forman bicapas na auga se está sometida a axitación. Esta estrutura é idéntica á das membranas celulares. Estas moléculas non se encontraban na Terra primixenia, aínda que outras cadeas anfifílicas longas tamén forman membranas. Ademais, estes corpos pódense expandir por inserción de lípidos adicionais, e baixo unha expansión excesiva poden sufrir escisións espontáneas que conservan o mesmo tamaño e composición de lípidos en ambas as proxenies. A idea principal desta teoría é que a composición molecular dos corpos lipídicos é a primeira forma de almacenar información e a evolución conduce á aparición de entidades poliméricas como o ARN ou el ADN que poden almacenar información favorabelmente. Aínda non se falou de ningún mecanismo que apoie a teoría do mundo de lípidos.
O modelo do polifosfato[editar | editar a fonte]
O problema con moitos dos modelos de abioxénese é que o equilibrio termodinámico favorece aos aminoácidos dispersos máis que aos seus polímeros, os polipéptidos; é dicir, que a polimerización é endotérmica. O que fai falta é unha causa que promova a polimerización. Unha solución ao problema pode encontrarse nas propiedades dos polifosfatos.[64][65] Os polifosfatos fórmanse pola polimerización dos ións ordinarios de monofosfato (PO4−3) baixo a acción da radiación ultravioleta. Os polifosfatos poden catalizar a polimerización dos aminoácidos a polipéptidos, reducindo a barreira de enerxía e facendo así factíbel o proceso.
Hipótese do mundo de HAP[editar | editar a fonte]
- Artigo principal: Hipótese do mundo de HAP.

Postuláronse outras fontes de moléculas complexas, incluíndo algunhas de orixe extraterrestre, estelar ou interestelar. Por exemplo, sábese a partir de análises espectrais e directos que hai moléculas orgánicas en cometas e meteoritos. En 2004 un equipo de investigación detectou trazas de hidrocarburos aromáticos policíclicos (HAPs) nunha nebulosa.[66] Estas son as moléculas máis complexas achadas no espazo até o momento. O uso dos HAP tamén foi proposto como precursor do mundo de ARN na hipótese do mundo de HAP[67]. O Telescopio Espacial Spitzer detectou recentemente unha estrela, a HH 46-IR, que está formándose nun proceso similar ao do Sol. No disco de material que rodea a estrela hai unha gran variedade de moléculas que inclúen os compostos de cianuro, hidrocarburos e hidróxido de carbono. Tamén se atoparon HAPs por toda a superficie da galaxia M81, que está a 12 millóns de anos luz da Terra, confirmando a súa ampla distribución no espazo.[68]
O modelo da ecopoese[editar | editar a fonte]
O modelo da ecopoese desenvolvérono os científicos brasileiros Félix de Sousa e Rodrigues Lima.[69] É un modelo que integra elementos e observacións de varios outros. Nesta teoría é o ambiente físico (οικόσ) o que promove a aparición da vida nos estadios temperáns en lugar de facelo na aparición ao azar de organismos que posteriormente condicionan o contorno, en especial en canto á acumulación de osíxeno. Propón que os ciclos xeoquímicos dos elementos bioxénicos, dirixidos por unha atmosfera primordial rica en osíxeno procedente da fotólise do vapor de auga evaporado dos océanos e hipercarbónica, puideron ser a base dun metabolismo planetario de carácter espacialmente continuo e global, que precedería e condicionaría a aparición gradual dunha vida como a actual, organizada en organismos descontinuos (individualizados).
Algunhas das súas predicións son as seguintes:
- A fotólise e posterior escape do hidróxeno acumularían unha cantidade significativa de osíxeno na atmosfera primitiva.
- Xeraríase un potencial redox entre as zonas de produción fotolítica de osíxeno e o ambiente submarino con minerais redutores, en especial ferro divalente.
- Estas interaccións darían lugar a un metabolismo global de base xeoquímica, o holoplasma, coa aparición de "ciclos" (como os actuais do carbono ou do fósforo) de elementos bioxénicos.
- Este "protometabolismo" debería ser congruente coas principais rotas metabólicas existentes hoxe en día.
- No medio hipercarbónico aumentan os catións divalentes e por iso a carboxilación (fixación de CO2 a outras moléculas) é enerxeticamente favorábel. Requiriríase, non obstante, de certos hidrocarburos da litosfera, en especial acetileno, que podería converterse por hidratación e carboxilación en intermediarios do ciclo de Krebs reversíbel. Este sería o elemento máis característico da circulación do carbono na ecosfera primitiva.
- Aplícase o principio de congruencia: postúlase a existencia dunha continuidade entre os factores protobiolóxicos ambientais e o metabolismo actual. Este pódese rastrexar nos coenzimas claves. Posteriormente produciríase a incorporación destas actividades a unidades catalíticas durante o mundo de ARN. O feito de que estes coenzimas estean relacionados estruturalmente cos nucleótidos con ribosa parece confirmar este extremo. O ambiente hipercarbónico tamén favorecería a propagación quiral dun dos enantiómeros unha vez seleccionado.
- O camiño cara ás protocélulas realizaríase mediante unha sucesión de "hábitos". En principio habería unha agregación de materia orgánica (hábito flocular) debida a principios sinxelos como a baixa actividade de auga, a coalescencia hidrófoba e a formación de tioácidos e a súa extracción parcial na fase lipídica.
- Na seguinte fase evolutiva (hábito reticular) aumenta a integración entre as fases dos flóculos (lipídica, peptídica e de polímeros fosforilados) dando lugar á tradución. Se agregarían vacúolos "metabólicos" con vacúolos ácidos ou respiratorios, dando lugar a retículos que posteriormente se fusionarían nun único compartimento co "hábito celular".
Hipótese da xénese múltipla[editar | editar a fonte]
Produciríase a aparición de diferentes formas de vida case simultaneamente na historia temperá da Terra.[70]. Dado que parece existir un único antepasado común entre todos os seres vivos, as outras formas estarían extinguidas, deixándonos fósiles a través da súa diferente bioquímica. Por exemplo, polo uso doutros elementos, como o arsénico en lugar do fósforo, e sobrevivindo como extremófilas ou simplemente permanecendo ignoradas ao seren análogas aos organismos actuais. Hartman, por exemplo, combina algunhas teorías propoñendo o seguinte:
Os primeiros organismos autorreplicantes foron arxilas ricas en ferro que fixaban dióxido de carbono no ácido oxálico e outros ácidos dicarboxílicos. O sistema de replicación das arxilas e o seu fenotipo metabólico evolucionou entón cara á rexión rica en sulfuro do manancial hidrotermal, adquirindo a capacidade de fixar nitróxeno. Finalmente incorporouse o fosfato no sistema en evolución que permitía a síntese de nucleótidos e fosfolípidos. Se a biosíntese recapitula a biopoese, entón a síntese dos aminoácidos precedeu á síntese de bases púricas e pirimidínicas. Máis alá diso a polimerización dos tioésteres de aminoácido en polipéptidos precedeu á polimerización dirixida de ésteres de aminoácidos por polinucleótidos.[71]
Argumentos contrarios á orixe abioxénica[editar | editar a fonte]
O moderno concepto de abioxénese sufriu as críticas dos científicos ao longo dos anos. O astrónomo Sir Fred Hoyle pronunciouse neste sentido baseándose na probabilidade de que a abioxénese suceda por azar. O físico Hubert Yockey criticaba a abioxénese no sentido de crela máis próxima á teoloxía que á ciencia. Outros científicos propuxeron contrapuntos á abioxénese, como Harold Urey, Stanley Miller, Francis Crick (biólogo molecular) e tamén cabería aliñar neste sentido a hipótese da panspermia dirixida de Leslie Orgel.
É difícil probar ou desmentir a abioxénese; por tanto, a hipótese ten moitas críticas, tanto da comunidade científica como desde posicións non científicas. Non obstante, a investigación e a xeración de hipóteses continúan coa esperanza de desenvolver un mecanismo teórico satisfactorio da abioxénese.
Hoyle[editar | editar a fonte]
Sir Fred Hoyle, xunto con Chandra Wickramasinghe, foi un crítico da abioxénese. En concreto, Hoyle rexeitaba que a evolución química puidera explicar a orixe natural da vida: o seu argumento baseábase principalmente na improbabilidade de que os que se estima que foron os compoñentes necesarios chegaran a agregarse pola evolución química. Aínda que as teorías modernas tratan este argumento, Hoyle nunca viu a evolución química como unha explicación razoábel. Hoyle prefería a panspermia como unha explicación natural alternativa da orixe da vida na Terra.
Yockey[editar | editar a fonte]
O teórico da información Hubert Yockey argumentaba que a investigación sobre a evolución química se enfronta aos seguintes problemas:
A investigación da orixe da vida parece ser única na conclusión de que foi xa aceptada como autorizada... o que aínda non se fixo foi encontrar os escenarios que describen o mecanismo detallado e os procesos polos cales sucedeu. Débese concluír que, contrariamente ao actual, establecido e sabio escenario que describe a xénese da vida na Terra por azar e causas naturais que poida ser aceptado con base nos feitos e non na fe, aínda non foi escrito.[72]
Nun libro que escribiu 15 anos despois, Yockey defendía a idea de que a abioxénese xurdiría a partir dun caldo primitivo é un paradigma errado:
Aínda que cando comezou o paradigma era digno de consideración, agora todo o esforzo empregado no primitivo paradigma do caldo tornouse en decepción na ideoloxía dos seus campións. (...) A historia da ciencia amosa que un paradigma, unha vez que adquiriu un status de aceptación (e se incorpora aos libros de texto) a pesar dos seus fallos, só se declara inválido cando se dispón doutro paradigma que ocupe o seu lugar. Non obstante, co obxecto de xerar progreso na ciencia, é necesario facer limpeza nos andeis, por así dicilo, de paradigmas falidos. Isto deberíase facer incluso se deixa os andeis completamente limpos e non sobrevive ningún paradigma. É unha característica do verdadeiro crente na relixión, filosofía e ideoloxía de que debe ter un conxunto de crenzas pase o que pase (Hoffer, 1951). A crenza nun caldo primitivo baseándonos en que non temos ningún outro paradigma é un exemplo da falacia lóxica da falsa alternativa. Na ciencia é unha virtude recoñecer a ignorancia. Este foi o caso universalmente na historia da ciencia, tal e como Kuhn (1970) discutiu en detalle. Non hai razón para que isto sexa diferente na investigación da orixe da vida. [73]
Yockey, en xeral, manifesta unha actitude altamente crítica cara aos que dan creto ás orixes naturais da vida, a miúdo facendo uso de palabras como fe ou ideoloxía. As publicacións de Yockey fixéronse as favoritas nas mans dos creacionistas, aínda que el non se considera a si mesmo como creacionista.[74]
Notas[editar | editar a fonte]
- ↑ NASA, ed. (2008). "Year Results on the Oldest Light in the Universe". Consultado o 27/01/2011. (en inglés)
- ↑ 2,0 2,1 WILDE, S.A.; VALLEY, J.W.; PECK, W.H.; GRAHAM, C.M. (11/01/2001). www.nature.com, ed. "Evidence from detrital zircons for the existence of continental crust and oceans on the Earth 4.4 Gyr ago". Consultado o 27/01/2011. (en inglés)
- ↑ TANZELLA-NITTI, G. www.disf.org, ed. "Dizionario Interdisciplinare di Scienza e Fede". Consultado o 28/01/2011. (en italiano)
- ↑ SHIELDS, Ch. (2009). Edward N. Zalta, ed. "Aristotle". Stanford Encyclopedia of Philosophy. Consultado o 27/01/2011. (en inglés)
- ↑ "Aristotle". Encyclopædia Britannica Online. Consultado o 27/01/2011. (en inglés)
- ↑ 6,0 6,1 6,2 FERNÁNDEZ RUIZ, B. (1980). La vida: origen y evolución. Barcelona, Salvat. (en castelán)
- ↑ Fernández Ruiz, B. (1980): La vida: origen y evolución. Barcelona: Salvat, páx. 7.
- ↑ Fernández Ruiz, B. (1980): La vida: origen y evolución. Barcelona: Salvat, páxs. 8-9.
- ↑ DREUX, P. (1974). Introducción a la ecología. Madrid, Alianza. (en castelán)
- ↑ KORMODY, E.J. (1973). Conceptos de ecología. Madrid, Alianza. (en castelán)
- ↑ PARRA, F. (1984). Diccionario de ecología, ecologismo y medio ambiente. Madrid, Alianza. (en castelán)
- ↑ FRIEDEL, H. (1980). Dicionário de ecologia e do meio ambiente. Porto, Lello & irmão. (en portugués)
- ↑ Fernández Ruiz, B. (1980): La vida: origen y evolución. Barcelona: Salvat, páxs. 14-15.
- ↑ Fernández Ruiz, B. (1980): La vida: origen y evolución. Barcelona: Salvat, páxs. 16-17.
- ↑ Fernández Ruiz, B. (1980): La vida: origen y evolución. Barcelona: Salvat, páxs. 18-19.
- ↑ "Life In a Soup – First Life on Earth". www.windmillministries.org (en inglés). Arquivado dende o orixinal o 29/06/2012. Consultado o 27/01/2011.
- ↑ NOWAK, M.A.; OHTSUKI, H. (14/07/2008). www.pnas.org, ed. "Prevolutionary dynamics and the origin of evolution" (PDF). Arquivado dende o orixinal (Pdf) o 09/06/2014. Consultado o 28/01/2011. (en inglés)
- ↑ ALBERTS, JOHNSON, LEWIS, RAFF, ROBERTS, WALTER (2002). Molecular Biology of the Cell. Routledge. ISBN 0-8153-3218-1. (en inglés)
- ↑ 19,0 19,1 Committee on Grand Research Questions in the Solid-Earth Sciences, National Research Council (2008). Origin and Evolution of Earth: Research Questions for a Changing Planet. The national academics press. ISBN 0-309-11886-7. (en inglés)
- ↑ CARO, G.; BOURDON, B.; BIRCK, J.L.; MOORBATH, S. (2003). "146Sm-142Nd evidence from Isua metamorphosed sediments for early differentiation of the Earth's mantle". Nature 423 (6938). PMID 12761546. (en inglés)
- ↑ CARLSON, R.W.; BOYET, M.; HORAN, M. (2007). "Chondrite barium, neodymium, and samarium isotopic heterogeneity and early Earth differentiation". Science 316 (5828). PMID 17525335. (en inglés)
- ↑ MORSE, J.W.; MacKENZIE, F.T. (1998). "Hadean Ocean Carbonate chemistry". Aquatic Geochemistry (4): 301–319.
- ↑ name=Neodimio>O'NEIL, J.; CARLSON, R.W.; FRANCIS, D.; STEVENSON, R.K. (2008). "Neodymium-142 Evidence for Hadean Mafic Crust". Science 321 (5897). DOI: 10.1126/science.1161925. (en inglés)
- ↑ SLEEP, N.H. et alt. (1989). "Annihilation of ecosystems by large asteroid impacts on early Earth". Nature (342): 139–142. (en inglés)
- ↑ 25,0 25,1 MOJKZSIS, S.J. et alt. (1996). "Evidence for life on earth before 3,800 million years ago". Nature (384): 55–59. (en inglés)
- ↑ LAZCANO, A.; MILLER, S.L. (1994). "How long did it take for life to begin and evolve to cyanobacteria". Journal of Molecular Evolution (39): 546–554. (en inglés)
- ↑ MAHER, K.A.; STEPHENSON, D.J. (18/02/1988). "Impact frustration of the origin of life". Nature 331: 612–614. (en inglés)
- ↑ ROBERTSON, M.P.; MILLER, S.L. (1995). "An Efficient Prebiotic Synthesis of Cytosine and Uracil". Nature (375): 772–774. (en inglés)
- ↑ BADA, J.L.; BIGHAM, C.; MILLER, S.L. (1994). "Impact Melting of Frozen Oceans on the Early Earth: Implications for the Origin of Life". Proceedings of the National Academy of Sciences: 1248–1250. (en inglés)
- ↑ LEVY, M.; MILLER, S.L.; BRINTON, K.; BADA, J.L. (2000). "Prebiotic synthesis of adenine and amino acids under Europa-like conditions". Icarus 145 (2): 609–613. PMID=11543508. Consultado o 28/01/2011. (en inglés)
- ↑ TRINKS, H.; SCHRÖDER, W.; BIEBRICHER, Ch. (2005). "Ice And The Origin Of Life". Origins of Life and Evolution of the Biosphere 35 (5): 429–445. doi=10.1007. Arquivado dende o orixinal o 12/01/2012. Consultado o 28/01/2011. (en inglés)
- ↑ FOX, D. (2008). "Did Life Evolve in Ice?". Discover Magazine. Consultado o 28/01/2011. (en inglés)
- ↑ OPARIN, A.I. (1968). The Origin and Development of Life. NASA TTF-488. Washington, D.C.L GPO. (en inglés)
- ↑ OPARIN, A.I. (1952). The Origin of Life. New York, Dover. (en inglés)
- ↑ RAULIN, F. (1994). La aparición de la vida. Barcelona, RBA Ediciones. ISBN. (en castelán)
- ↑ BRYSON, B. (2003). A Short History of Nearly Everything. ISBN 0-552-99704-8. (en castelán)
- ↑ www.panspermia-theory.com (ed.). "Panspermia and the Origin of Life on Earth". Arquivado dende o orixinal o 03/02/2011. Consultado o 28/01/2011. (en inglés)
- ↑ YAMAGISHI, A.; ITAHASHI, S.; KOBAYASHI, H.; YOKOBORI, S., MOTONO, C. www4.jsforum.or.jp, ed. "Distribution of extremophiles in space environment". Arquivado dende o orixinal o 04/06/2009. Consultado o 28/01/2011. (en inglés)
- ↑ MARTINS, Z., et alt. (2008). "Extraterrestrial nucleobases in the Murchison meteorite". Earth and Planetary Science Letters 270 (1-2): 130–136. doi=10.1016/j.epsl.2008.03.026. Arquivado dende o orixinal o 19/02/2010. Consultado o 28/01/2011. (en inglés)
- ↑ BERALDI, H. Instituto de Geología (UNAM), ed. "Estromatolitos". Consultado o 28/01/2011. (en castelán)
- ↑ EMBLEY, T.M.; MARTIN, W. (2006). "Eukaryotic evolution, changes and challenges" (PDF). Nature Reviews 440 (30). doi:10.1038/nature04546. Arquivado dende o orixinal (PDF) o 07/10/2007. Consultado o 28/01/2011. (en inglés)
- ↑ POOLE, A.; PENNY, D. (2007). "Evaluating hypotheses for the origin of eukaryotes" (PDF). Bioessays 29 (1): 74–84. pmid=17187354. Arquivado dende o orixinal (PDF) o 13/01/2012. Consultado o 28/01/2011. (en inglés)
- ↑ Wächtershäuser, Günter (1992-01). "Groundworks for an evolutionary biochemistry: The iron-sulphur world". Progress in Biophysics and Molecular Biology (en inglés) 58 (2): 85–201. doi:10.1016/0079-6107(92)90022-X.
- ↑ HUBER, C.; WÄCHTERHÄUSER, G. (1998). "Peptides by activation of amino acids with CO on (Ni,Fe)S surfaces: implications for the origin of life". Science (281): 670–672. (en inglés)
- ↑ 45,0 45,1 MARTIN, W.; RUSSELL, M.J. (2002). "On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells". Philosophical Transactions of the Royal Society: Biological sciences 358 (1429): 59–85. doi=10.1098/rstb.2002.1183. (en inglés)
- ↑ DARTNELL, L. (2008). "Life's a beach on planet Earth". New Scientist. (en inglés)
- ↑ ADAM, Z. (2007). "Actinides and Life's Origins". Astrobiology 7 (6): 852–872. doi=10.1089/ast.2006.0066; pmid=18163867. (en inglés)
- ↑ NOYES, H.; PIERRE, H.; BONNER, W.A.; TOMLIN, J.A. (1977). "On the origin of biological chirality via natural beta-decay". Origins of Life and Evolution of Biospheres 8 (1). (en inglés)
- ↑ HAZEN, R.M. (2005). Genesis: the scientific quest for life's origin. JosephHenry Books. (en inglés)
- ↑ CLARK, S. (1999). "Polarised starlight and the handedness of Life". American Scientist 97: 336–343. (en inglés)
- ↑ MULLEN, L. (09/05/2005). www.astrobio.net, ed. "Building Life from Star-Stuff". Arquivado dende o orixinal o 27/03/2014. Consultado o 28/01/2011. (en inglés)
- ↑ GILBERT, W. (1986). "The RNA World" 319. doi=10.1038/319618a0. (en inglés)
- ↑ JOHNSTON, W.K.; UNRAU, P.J.; LAWRENCE, M.S.; GLASNER, M.E.; BARTEL, D.P. (2001). "RNA-Catalyzed RNA Polymerization: Accurate and General RNA-Templated Primer Extension". Science 292 (1319). Consultado o 28/01/2011. (en inglés)
- ↑ ORGEL, L. (2000). "A Simpler Nucleic Acid". Science 290 (5495): 1306 – 1307. (en inglés)
- ↑ NELSON, K.E.; LEVY, M.; MILLER, S.L. (2000). "Peptide nucleic acids rather than RNA may have been the first genetic molecule". Proc. Natl. Acad. Sci. USA (97): 3868–3871. Arquivado dende o orixinal o 25/02/2008. Consultado o 28/01/2011. (en inglés)
- ↑ ROBINSON, R.; coautores, se os houber (2005). "Jump-Starting a Cellular World: Investigating the Origin of Life, from Soup to Networks". PLoS 3 (11). doi=10.1371/journal.pbio.0030396. Consultado o 28/01/2011. (en inglés)
- ↑ PNAS, vol. 97, no. 23, Novembro 7 2000, p12503-12507
- ↑ Esta teoría está ampliada no libro A célula: Evolución do primeiro organismo de Joseph Panno Ph.D.
- ↑ GRIFFITH, R.W. (2009). "A Specific Scenario for the Origin of Life and the Genetic Code Based on Peptide/Oligonucleotide Interdependence". Orig Life Evol Biosph. pmid =19468859. (en inglés)
- ↑ Genetic Takeover: And the Mineral Origins of Life. ISBN 0-521-23312-7. (en inglés)
- ↑ BULLARD, T.; FREUDENTHAL, J.; AVAGYAN, S.; KAHR, B. (2007). "Test of Cairns-Smiths crystals-as-genes hypothesis". Faraday Discuss. DOI: 10.1039/b616612c. (en inglés)
- ↑ MOORE, C. (2007). "Crystals as genes?". Chemical Science. Consultado o 28/01/2011. (en inglés)
- ↑ AA.VV. ool.weizmann.ac.il, ed. "Origin Of Life research". Arquivado dende o orixinal o 10/07/2007. Consultado o 28/01/2011. (en inglés)
- ↑ BROWN, M.R.W.; KORNBERG, A. (2004). "Inorganic polyphosphate in the origin and survival of species". Proc. Natl. Acad. Sci. USA. 101 (46): 16085–16087. doi=10.1073/pnas.0406909101. Consultado o 28/01/2011. (en inglés)
- ↑ CLARK, D.P. www.science.siu.edu, ed. "The origin of life". Arquivado dende o orixinal o 21/07/2011. Consultado o 28/01/2011. (en inglés)
- ↑ WITT, A.N.; VIJH, U.P.; GORDON, K.D. www.aas.org, ed. "Discovery of Blue Fluorescence by Polycyclic Aromatic Hydrocarbon Molecules in the Red Rectangle". Arquivado dende o orixinal o 14/02/2008. Consultado o 28/01/2011. (en inglés)
- ↑ BATTERSBY, S. (2004). Newscientist, ed. "Space molecules point to organic origins". Consultado o 28/01/2011. (en inglés)
- ↑ MULLEN, L. (23/12/2003). "First Images Show Organic Molecules". Consultado o 28/01/2011. (en inglés)
- ↑ FÉLIX DE SOUSA, R.A. (2006). A criação da ecosfera. Ecopoese, 2ª Ed., Rio de Janeiro. (en portugués)
- ↑ DAVIES, P. (2007). "Are Aliens Among Us?". Scientific American. Consultado o 28/01/2011. (require subscrición (?)). (en inglés)
- ↑ HARTMAN, H. (1998). "Photosynthesis and the Origin of Life". Origins of Life and Evolution of Biospheres 28 (4–6.). (en inglés)
- ↑ YOCKEY (1977). "A calculation of the probability of spontaneous biogenesis by information theory". Journal of Theoretical Biology (67): 377–398. (en inglés)
- ↑ YOCKEY (1992). Information Theory and Molecular Biology. Cambridge University Press. ISBN 0-521-80293-8. (en inglés)
- ↑ YOCKEY (26/02/1996). www.asa3.org, ed. "Yockey#9". Arquivado dende o orixinal o 20/12/2011. Consultado o 28/01/2011. (en inglés)
Véxase tamén[editar | editar a fonte]
Bibliografía[editar | editar a fonte]
- Brooks, J. e Shaw, G. (1973): Origins and Development of Living Systems. Academic Press. ISBN 0-12-135740-6
- Arnau, C. e Carbó, E. (1974): El origen de la vida. Barcelona: Salvat. ISBN 84-345-7375-X Os autores fan un repaso ao problerma da vida, o escenario da mesma (no universo), as principais hipóteses sobre a orixe da vida, a síntese das moléculas prebióticas, a orixe da clave xenética, e a exobioloxía. O libro inclúe unha entrevista con Joan Oró.
- De Duve, C. (1966): Vital Dust: The Origin and Evolution of Life on Earth. Basic Books. ISBN 0-465-09045-1
- Dreux, P. (1975): Introducción a la ecología. Madrid: Alianza Editorial. ISBN 84-206-1586-2
- Fernández Ruiz, B. (1980): La vida: origen y evolución. Barcelona: Salvat. ISBN 84-345-7807-7
- Grobstein, C. (1973): La estrategia de la vida. Barcelona: Blume. ISBN 84-345-7807-7 Neste libro o autor non insiste tanto na orixe da vida como na complexidade da mesma, estudando aspectos tan interesantes como os niveis de organización, variación, diversidade etc. En todo el queda de manifesto unha visión evolutiva.
- Horgan, J. (1991): "In the beginning". Scientific American, Volume 264, páxs. 100-109.
- Huber, C. and Wächterhäuser, G. (1998): "Peptides by activation of amino acids with CO on (Ni, Fe) S surfaces: implications for the origin of life". Science, Volume 281, páxs. 670–672.
- Luisi, P. L. (2010): La vida emergente: de los orígenes químicos a la biología sintética. Barcelona: Tusquets. Metatemas. ISBN 978-84-8383-204-2
- Martin, W. e Russell, M. J. (2002): "On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells". Philosophical Transactions of the Royal Society: Biological sciences, Volume 358, páxs. 59-85.
- Keosian, J. (1975): El origen de la vida. Madrid: Alhambra. Este libro expón fundamentalmente o problema da orixe química da vida, sen tratar a evolución posterior. Analiza o concepto de vida e as concepcións de distintas escolas. Á final de cada capítulo achega abundante bibliografía.
- Maynard Smith, J. e Szathmary, E. (2000): The Origins of Life: From the Birth of Life to the Origin of Language. Oxford Paperbacks. ISBN 0-19-286209-X
- Dyson, F. J., (1999): Los orígenes de la vida. Cambridge University Press. ISBN 978-84-8323-097-8. O físico e comunicador F. J. Dyson ofrece modelos alternativos para a orixe dos primeiros sistemas viventes. Non é un resume do coñecemento sobre o tema, senón unha perspectiva excéntrica pero ben fundada e moi suxerente.
- Raulin, F. (1994): La aparición de la vida. Barcelona: RBA Editores. ISBN 84-473-0283-0
- Schrödinger, E. (1984): ¿Qué es la vida?. Barcelona: Tusquets. ISBN 978-84-7223-607-3
- Seifert, J. (1997): What is Life. The Originality, Irreducibility and Value of Life. Amsterdam: Value Inquiry Books Series, Rodopi.
- Oparin, A. (1979): El origen de la vida. Madrid: Akal. Neste libriño o autor expón a súa coñecida teoría acerca da evolución química desde a formación de hidrocarburos sinxelos até as máis complexas macromoléculas. Expón as súas ideas sobre a formación de coacervados como paso intermedio cara a constitución dun ser vivo primitivo.
- Haczyc, M. M. e Szostak, J. W. (2004): "Replicating vesicles as models of primitive cell growth and division". Current Opinion in Chemical Biology, 8, páxs. 660–664.[1]
- Hazen, R. M. (2005): Genesis: The Scientific Quest for Life's Origins. Joseph Henry Press. ISBN 0-309-09432-1
- Cairns Smith: originsoflife.net: ilustración dunha posíbel solución usando o comportamento de cristais de arxilas
- Deep Space: Scientists Find Clues That Life Began NASA Astrobiology Institute.
- Gold, T.: The Deep Hot Biosphere Theory
- Orgel, L.: Self-organizing biochemical cycles Arquivado 29 de febreiro de 2008 en Wayback Machine.
- PNAS: Modelo da orixe da vida que implica as ceolitas [2]
- Schopf, J. W. et al. (2002): "Laser-Raman imagery of Earth's earliest fossils". Nature, Volume 416, páxs. 73-76. PMID 11882894
Outros artigos[editar | editar a fonte]
- Evolución biolóxica
- Bioxénese
- Historia da Terra
- Historia evolutiva da vida
- Astrobioloxía
- Hipótese da Terra púrpura
- Primeiro antepasado común universal (FUCA)
- Último antepasado común universal (LUCA)
Ligazóns externas[editar | editar a fonte]
- Web da Sociedade Internacional para o Estudo da Orixe da Vida (en inglés).
- Vídeo en inglés 'A orixe da vida, por John Maynard-Smith' Discurso perante a Real Instituciónn pola sociedade Vega Science
- Web sobre a orixe da vida que inclúe artigos e recursos do Dr. Michael Russell, da Universidade de Glasgow
- Possible Connections Between Interstellar Chemistry and the Origin of Life on the Earth
- It's alive - isn't it?
- How Life Began: New Research Suggests Simple Approach
| Evolución |  | |
|---|---|---|
| Xenética de poboacións | ||
| Desenvolvemento | ||
| Tempo e modos | ||
| Especiación | ||
| Historia | ||