
Coleópteros
| Coleópteros Rango fósil: 299 Ma-0 Ma Permiano-Actual | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 De esquerda a dereita: Lamprima aurata, Megasoma, Amblytelus, Chondropyga dorsalis e Rhinotia hemistictus. | |||||||||||||||
| Clasificación científica | |||||||||||||||
| |||||||||||||||
| Subordes | |||||||||||||||
Os coleópteros[2] (Coleoptera) son un grupo de insectos que constitúen unha orde dentro da superorde Endopterygota. Moitos deles reciben o nome de escaravellos[3], que se usa ás veces como termo xeral,[4] pero outros teñen outros nomes específicos como gurgullos, vagalumes, couzas da madeira, maruxiñas, vacalouras... O seu par de ás anterior, chamadas élitros, está endurecido formando unha especie de estoxo, o que os distingue da maioría dos outros insectos. Os Coleoptera, cunhas 400 000 especies, son a maior de todas as ordes de insectos, constitúen case o 40% dos insectos descritos e o 25% de todos os animais coñecidos; descóbrense novas especies decote. A máis grande de todas as familias é a dos Curculionidae (gurgullos) cunhas 70 000 especies. Encóntranse en case todos os hábitats excepto no mar e as rexións polares. Interaccionan cos seus ecosistemas de varios modos, xa que hai especies herbívoras, detritívoras e carnívoras: aliméntanse de plantas e fungos, de restos de animais e plantas descompostos, ou comen outros invertebrados. Algunhas especies son graves pragas agrícolas, como o escaravello da pataca ou larvas dalgunhas especies, mentres que outros, como os Coccinellidae (xoaniñas ou maruxiñas) comen áfidos, cochinillas, trips e outros insectos que zugan o zume das plantas ou danan doutros modos as colleitas.
Os coleópteros teñen un exoesqueleto especialmente duro, incluíndo os élitros, aínda que algúns, como os estafilínidos teñen élitros moi curtos, e os meloidos teñen élitros brandos. A anatomía xeral dun coleóptero é bastante uniforme e típica dos insectos, aínda que hai varios exemplos novidosos, como as adaptacións que se observan nos escaravellos acuáticos, que atrapan burbullas de aire baixo os élitros para usalas mentres están mergullados. Os coleópteros son endopterigotos, o cal significa que sofren unha metamorfose completa, experimentando unha serie de cambios ben aparentes e relativamente abruptos na estrutura corporal entre a eclosión do ovo e a etapa adulta, despois de pasaren por unha fase pupal relativamente inmóbil. Algúns, como os lucánidos (por exemplo as vacalouras), teñen un marcado dimorfismo sexual, os machos posúen mandíbulas enormemente agrandadas que utilizan para loitar con outros machos. Moitos coleópteros son aposemáticos, polo que teñen cores rechamantes e padróns de coloración de advertencia da súa toxicidade, mentres que outros son imitadores batesianos inofensivos de ditos insectos. Moitos outros coleópteros, incluíndo os que viven en lugares areosos, teñen camuflaxes efectivas.
Os escaravellos son insectos moi importantes na cultura humana, desde os escarabeos sagrados do Antigo Exipto á antiga arte asiática de ás de escaravellos (utiliza ás de escaravellos iridescentes para decorar) e o uso como mascotas ou na loita de insectos para o entretemento e apostar. Moitos grupos de coleópteros teñen coloracións vivas e atractivas, polo que son coleccionados ou usados con fins decorativos. Unhas 300 especies son utilizadas como alimento humano, especialmente as larvas; entre as especies máis consumidas están as larvas do escaravello da fariña e do escaravello rinoceronte. Porén, o maior impacto dos coleópteros na vida humana é na agricultura, explotación forestal e horticultura, xa que algúns son importantes pragas. Entre as pragas máis graves están as do gurgullo do algodoeiro, o escaravello da pataca, o escaravello do coqueiro e o escaravello do piñeiro. Porén, a maioría dos coleópteros, non causan danos económicos e moitos, como as xoaniñas (predadoras) e escaravellos peloteiros (que se alimentan de feces), son beneficiosos e axudan a controlar as pragas de insectos ou son bos para os ecosistemas.
Etimoloxía[editar | editar a fonte]

O nome Coleoptera procede do grego koleopteros (κολεόπτερος), que lle deu a este grupo Aristóteles debido aos seus duros élitros, procedente de koleos, 'vaíña, estoxo' e pteron, 'á'.[5] Escaravello procede do latín scarafaius que derivou de scarabaeus, 'escaravello',[4] e este á súa vez procede do grego karabos ou skarabeios, 'escaravello'.[6]
Distribución e diversidade[editar | editar a fonte]
Os coleópteros son con gran difeenza a orde máis grande de insectos: as súas aproximadamente 400 000 especies supoñen o 40% de todas as especies de insectos descritas e un 25% de todos os animais.[1][7][8][9][10][11] Un estudo de 2015 ofreceu catro estimacións independentes do número total de especies de coleópteros que podería haber, dando unha estimación media dunhas 1,5 millóns cun "rango sorprendentemente estreito"[12] e as catro estimacións ían desde un mínimo de 0,9 a un máximo de 2,1 millóns de especies. As catro estimacións utilizaron relacións de especificidade de hóspede (de 1,5 a 1,9 millóns), proporcións con outros taxons (de 0,9 a 1,2 millóns), proporcións plantas: escaravellos (de 1,2 a 1,3), e extrapolacións baseadas no tamaño corporal por ano de descrición (de 1,7 a 2,1 millóns).[12][13]
Os coleópteros encóntranse en case todos os hábitats, incluíndo as augas docese os hábitats costeiros, en calquera sitio onde haxa vexetación, desde árbores e as súas codias a flores, follas e baixo terra preto das raíces, incluso dentro das plantas en bugallas, en todos os tecidos das plantas, incluíndo os mortos ou podres.[14]
O escaravello máis pesado, e tamén o insecto máis pesado, é a larva do escaravello goliat, Goliathus goliatus, que pode alcadar unha masa de polo menos 115 g e unha lonxitude de 11,5 cm. Os machos adultos do escaravello goliat son os coleópteros máis pesados en estado adulto, pesando de 70 a 100 g e medindo ata 11 cm.[15] Os escaravellos elefante adultos, Megasoma elephas e Megasoma actaeon a miúdo chegan a 50 g e 10 cm.[16]
O coleóptero máis longo é o escaravello hércules, Dynastes hercules, que ten unha lonxitude máxima global de polo menos 16,7 cm incluíndo o seu longo corno pronotal. O coleóptero máis pequeno rexistrado (en 2015) e tamén o insecto de vida libre máis pequeno, é o Scydosella musawasensis, que mide só 325 µm de lonxitude.[17]
-
 O escaravello titán, Titanus giganteus, un cerambícido tropical, é un dos insectos máis grandes e pesados do mundo.
O escaravello titán, Titanus giganteus, un cerambícido tropical, é un dos insectos máis grandes e pesados do mundo. -
 Scydosella musawasensis, o escaravello máis pequeno coñecido: a barra de escala da dereita é de 50 micrómetros.
Scydosella musawasensis, o escaravello máis pequeno coñecido: a barra de escala da dereita é de 50 micrómetros. -
 O escaravello hércules, Dynastes hercules ecuatorianus, é o máis longo de todos os escaravellos
O escaravello hércules, Dynastes hercules ecuatorianus, é o máis longo de todos os escaravellos -
 Un Protaetia cuprea iridiscente alimentándose sobre un cardo
Un Protaetia cuprea iridiscente alimentándose sobre un cardo




Evolución[editar | editar a fonte]
Paleozoico tardío e Triásico[editar | editar a fonte]


O insecto fósil máis antigo que inequivocamente lembra un coleóptero data do Permiano inferior, hai uns 270 millóns de anos, aínda que estes espécimes, pertencentes á familia Tshekardocoleidae, teñen antenas de 13 segmentos, élitros cunha venación máis desenvolvida e unhas costelas ou nervaduras lonxitudinais máis irregulares, e o abdome e o ovipositor estendíanse alén do ápice dos élitros. Os primeiros escaravellos do Permiano, que se agrupan no taxon "Protocoleoptera", pénsase que eran xilófagos e perforaban a madeira. No evento de extinción do Permiano–Triásico acontecido a finais do Permiano, extinguíronse un 30% de todas as especies de insectos, polo que o rexistro fósil de insectos só inclúe escaravellos desde o Triásico inferior, hai 220 millóns de anos. Por esa época, durante o Triásico tardío, apareceron as especies comedoras de fungos como os Cupedidae no rexistro fósil. No Triásico superior empezaron a aparcer insectos comedores de algas como os Triaplidae e Hydrophilidae, xunto cos escaravello acuáticos predadores. Os primeiros gurgullos, incluíndo os Obrienidae, apareceron xunto aos primeiros Staphylinidae, que lembran moito ás especies recentes.[18] Algúns entomólogos son excépticos de que eses insectos tan antigos estean estreitamente emparentados coas especies actuais, argumentando que isto é extremadamente improbable; por exemplo, a estrutura do metepisterno suxire que os Obrienidae poderían ser Archostemata, que non son gurgullos, malia que os fósiles teñen un morro similar ao dos gurgullos.[19]
En 2009, describiuse un escaravello fósil do Pensilvaniano procedente de Mazon Creek, Illinois, que fai remontar a orixe dos coleópteros a unha data máis temperá, hai entre 318 e 299 millóns de anos.[20] Fósiles desa idade atopáronse en Asia e Europa, por exemplo nos estratos fosilíferos de lousa vermella de Niedermoschel preto de Mainz, Alemaña.[21] Un fósil alemán de algo máis de 295 millóns de anos de antigüidade é outro candidato a ser o escaravello máis antigo coñecido, que se clasificou no xénero Coleopsis do Permiano temperán (Asseliense).[22] Atopáronse máis fósiles en Obora, República Checa e Tshekarda nos Montes Urais, Rusia.[23] Porén, hai só uns poucos fósiles de Norteamérica antes do Permiano medio, aínda que tanto Asia coma Norteamérica se uniran a Euramérica na paleoxeografía da época. Os primeiros descubrimentos en Norteamérica, feitos na formación Wellington de Oklahoma, foron publicados en 2005 e 2008.[18][24]
Como consecuencia do evento de extinción do Permiano–Triásico, o rexistro fósil de insectos é escaso, incluíndo os escaravellos desde o Triásico inferior.[25] Con todo, hai unhas poucas excepcións, como en Europa do leste. No xacemento de Babiy Kamen na cunca de Kuznetsk, descubríronse numerosos fósiles de escaravellos, incluíndo espécimes enteiros das infraordes Archostemata (por exemplo, Ademosynidae, Schizocoleidae), Adephaga (por exemplo, Triaplidae, Trachypachidae) e Polyphaga (por exemplo, Hydrophilidae, Byrrhidae, Elateroidea).[26] Porén, as especies das familias Cupedidae e Schizophoroidae non están presentes neste xacemento, mentres que son as que predominan noutros xacementos fósiles desde o Triásico inferior, como en Khey-Yaga, Rusia, na cunca de Korotaikha.[18]
Xurásico[editar | editar a fonte]
Durante o Xurásico (desde hai 210 a 145 millóns de anos), houbo un drástico incemento na diversidade de familias de coleópteros,[18] incluíndo o desenvolvemento e crecemento de especies carnívoras e herbívoras. Os Chrysomeloidea diversificáronse aproximadmente nese momento, alimentándose dunha ampla gama de plantas hóspede desde cícadas e coníferas a anxiospermas.[27] Preto do Xurásico superior, os Cupedidae diminuíron, pero a diversidade das especies iniciais comedoras de plantas aumentou. A maioría dos coleópteros comedores de plantas recentes alimentábanse de plantas con flor anxiospermas, cuxo éxito contribuíu a duplicar o número de especies comedoras de plantas durante o Xurásico medio. Porén, o incremento do número de familias de coleópteros durante o Cretáceo non se correlaciona co incremento do número de especies de anxiospermas.[28] Nesa época, apareceron numerosos gurgullos primitivos (por exemplo os Curculionoidea) e escaravellos como por exemplo os Elateroidea. Os primeiros Buprestidae está presentes, pero seguiron sendo raros ata o Cretáceo.[29][30][31] Os primeiros Scarabaeidae non eran coprófagos senón que probablemente se alimentaban de madeira en descomposición con axuda dos fungos; son un exemplo antigo de relación mutualista.
Hai máis de 150 xacementos fósiles importantes do Xurásico, a maioría no leste de Europa e norte de Asia. Os máis salientables son: Solnhofen en Baviera, Alemaña,[32] Karatau no sur de Casaquistán,[33] a formación Yixian en Liaoning, norte da China,[34] e a formación Jiulongshan e outros xacementos de Mongolia. En Norteamérica só hai uns poucos xacementos con rexistros fósiles de insectos do Xurásico, concretamente nos depósitos de calcarias cunchíferas na cunca de Hartford, a cunca de Deerfield e a cunca de Newark.[18][35]
Cretáceo[editar | editar a fonte]
No Cretáceo produciuse a fragmentación da masa continental do sur (Gondwana), coa abertura do Atlántico sur e o illamento de Nova Zelandia, mentres que Suramérica, a Antártida e Australia se afastaron máis.[27] A diversidade de Cupedidae e Archostemata diminuíu considerablemente. Os escravellos de terra predadores (Carabidae) e os Staphylinidae empezaron a distribuírse en diferentes padróns; os Carabidae vivían predominantemente nas rexións cálidas, mentres que os Staphylinidae e Elateridae preferían climas temperados. Igualmente, as especies predadoras das superfamilias Cleroidea e Cucujoidea cazaban as súas presas baixo a codia das árbores xunto cos Buprestidae. A diversidade de bupréstidos incrementouse rapidamente, xa que eran os consumidores primarios de madeira,[36] mentres que os Cerambycidae eran bastante raros: a súa diversidade só aumentou contra o final do Cretáceo superior.[18] Os primeiros escaravellos coprófagos son do Cretáceo superior[37] e pode que viviran alimentándose dos excrementos dos dinosauros herbívoros.[38] Encóntranse agora as primeiras especies nas que as larvas e adultos están adaptados a un estilo de vida acuático. Os escaravellos Gyrinidae tiñan unha diversidade moderada, aínda que outros escaravellos iniciais dese tipo (por exemplo os Dytiscidae) eran menos diversos, e as máis estendidas eran as especies de Coptoclavidae, que depredaban larvas de moscas acuáticas.[18]
Moitos xacementos fósiles de todo o mundo conteñen coleópteros fósiles do Cretáceo. A maioría están en Europa e Asia e pertencen á zona de clima temperado durante o Cretáceo.[34] Entre os xacementos do Cretáceo inferior están os estratos fósiles de Crato na cunca de Araripe en Ceará, norte do Brasil, e tamén na formación Santana supraxacente; esta última estaba preto do ecuador daquela. En España, hai xacementos importantes preto de Montsec e Las Hoyas. En Australia, os estratos fosilíferos de Koonwarra do grupo Korumburra, South Gippsland, Victoria, son notables. Xacementos importantes do Cretáceo superior son os de Kzyl-Dzhar no sur de Casaquistán e Arkagala en Rusia.[18]
Cenozoico[editar | editar a fonte]

Os fósiles de coleópteros son abundantes no Cenozoico; no Cuaternario (iniciado hai 1,6 millóns de anos), as especies fósiles eran xa idénticas ás que viven hoxe, mentres que se nos remontamos ao Mioceno tardío (hai 5,7 millóns de anos) os fósiles son aínda tan próximos ás formas modernas que son probablemente os antepasados das especies vivas. As grandes oscilacións climáticas que tiveron lugar no Cuaternario causaron un cambio na distribución xeográfica dos coleópteros tan grande que a localización actual proporciona poucas pistas da historia bioxeográfica destas especies. É evidente que o illamento xeográfico das poboacións debeu romperse a miúdo a medida que os insectos se movían baixo a influencia do clima cambiante, causando unha mestura das pozas ou acervos xenéticos, evolución rápida e extincións, especialmente nas latitudes medias.[40]
Filoxenia[editar | editar a fonte]
O gran número de especies de coleópteros supuxo especiais problemas para a súa clasificación. Algunhas familias conteñen decenas de miles de especies e deben ser divididas en subfamilias e tribos. Este inmenso número fixo que o biólogo evolutivo J. B. S. Haldane bromease, cando algúns teólogos lle preguntaron que se podía inferir sobre a mente do Creador a partir das obras da Súa Creación, contestando "Un desorbitante cariño polos escaravellos".[41] A suborde maior é a Polyphaga, que contén máis de 300 000 especies descritas en máis de 170 familias, como a Staphylinidae, Scarabaeidae), Meloidae, Lucanidae e Curculionidae.[10][42] Estes grupos de coleópteros polífagos poden identificarse pola presenza de escleritos cervicais (partes endurecidas da cabeza usadas como puntos de unión dos músculos) ausentes nas outras subordes.[43] A suborde Adephaga contén unhas 10 familias de escaravellos principalmente predadores, incluíndo os Carabidae, os Dytiscidae e Gyrinidae. Nestes insectos, os testículos son tubulares e o primeiro esterno abdominal (unha placa do exoesqueleto) está dividida polas coxas traseiras (as articulacions basais das patas).[44] Archostemata contén catro familias de especies principalmente comedoras de madeira, incluíndo os Cupedidae e o Micromalthus debilis.[45] Os Archostemata teñen unha placa exposta chamada a metatrocantina en fronte do segmento basal ou coxa da pata traseira.[46] Os Myxophaga comprenden unhas 65 especies descritas distribuídas en catro familias, normalmente moi pequenas, como Hydroscaphidae e o xénero Sphaerius.[47] Os mixófagos son pequenos e principalmente comedores de algas. As súas pezas bucais teñen a característica de carecer de galeas e ter un dente móbil na mandíbula esquerda.[48]
A consistencia da morfoloxía dos coleópteros, en especial a súa posesión de élitros, indica que os Coleoptera son monofiléticos, mais hai dúbidas sobre a disposición das subordes, principalmente os Adephaga, Archostemata, Myxophaga e Polyphaga dentro dese clado.[27][49][50][51][52] Os insectos parasitos Strepsiptera crese que son un grupo irmán dos coleópteros, que se separou deles no Permiano temperán.[51][53][54][55]
A análise xenética filoxenética confirma que os Coleoptera son monofiléticos. Duane McKenna et al. (2015) utilizaron oito xenes nucleares de 367 especies de 172 das 183 familias de coleópteros. Os autores dividiron os Adephaga en dous clados, Hydradephaga e Geadephaga, fraccionaron os Cucujoidea en tres clados, e situaron os Lymexyloidea dentro de Tenebrionoidea. Os Polyphaga parecen datar do Triásico. A maioría das familias existentes de coleópteros parece que se orixinaron no Cretáceo.[55] O cladograma baséase en McKenna (2015).[55] O número de especies de cada grupo (principalmente superfamilias) móstrase entre paréntesess, e están en letra grosa se son máis de 10 000. As datas de orixe dos grupos maiores móstranse en cursiva en millóns de anos antes do presente (m. a.).[56]
| Coleoptera 285 m. a. |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||




Morfoloxía externa[editar | editar a fonte]

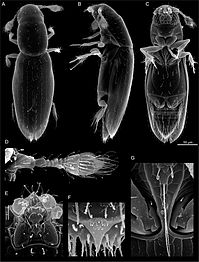
Os coleópteros caracterízanse de forma xeral por ter un exoesqueleto especialmente duro e duras ás anteriores (élitros) que non poden usar para voar. Case todos os coleópteros teñen mandíbulas que se moven nun plano horizontal. As pezas bucais raramente son suctoras, aínda que ás veces están reducidas; as maxilas sempre levan palpos. As antenas xeralmente teñen 11 segmentos ou menos, excepto nalgúns grupos como os cerambícidos e ripicéridos. As coxas das patas están normalmente situadas metidas dentro dunha cavidade coxal. As estruturas xenitais están pregadas no último segmento abdominal en todos os escaravellos existentes. As larvas poden ser confundidas con frecuencia coas doutros insectos endopterigotos.[46] O exoesqueleto dos coleópteros consta de numerosas placas chamadas escleritos, separadas por estruturas finas. Este deseño proporciona defensas acoirazadas de armadura á vez que se mantén a flexibilidade. A anatomía xeral dun coleóptero é bastante uniforme, aínda que os órganos específicos e os apéndices varían moito en aparencia e función entre as moitas familias da orde. Como todos os insectos, os corpos dos coleópteros están divididos en tres seccións: a cabeza, o tórax e o abdome.[8] Como hai tantas especies, a identificación precisa é bastante difícil e depende de atributos como a forma das antenas, a fórmula tarsal[a] e a forma dos pequenos segmentos das patas, as pezas bucais e as placas ventrais (esternos, pleuras, coxas). En moitas especies a identificación exacta só pode facerse examinando as características estruturas xenitais masculinas.[57]
Cabeza[editar | editar a fonte]
.jpg)
A cabeza ten as pezas bucais proxectadas cara a adiante ou ás veces voltadas cara a abaixo, está normalmente moi esclerotizada e ás veces é moi grande.[7] Os ollos son compostos e poden mostrar unha gran adaptabilidade, como no caso dos escaravellos acuáticos Gyrinidae, nos que están divididos para pemitir a visión por riba e por debaixo da tona da auga. Os cerambícidos e gurgullos teñen ollos divididos, mentres que moitos teñen ollos con amosega, e uns poucos teñen ocelos, que son ollos pequenos simples xeralmente situados máis atrás na cabeza (no vértex); estes son máis comúns nas larvas que nos adultos.[58] Os ocelos encóntranse en adultos de Dermestidae, nalgúns Omaliinae e nos Derodontidae.[58]

As antenas dos coleópteros son principalmente órganos para a percepción sensorial e poden detectar o movemento, olor e substancias químicas,[59] pero poden tamén usarse para sentir fisicamente (tacto) o ambiente no que vive. As distintas familias de coleópteros poden usar as antenas de diferentes maneiras. Por exemplo, cando se moven rapidamente, os escaravellos Cicindelinae non poden ver moi ben e o que fan é manter as antenas rixidas e por diante para evitar os obstáculos.[60] Certos cerambícidos usan as antenas para equilibrarse, e os escaravellos meloidos poden usalas para agarrar. Algúns escaravellos acuáticos poden usar as antenas para recoller aire e traspasalo baixo o seu corpo mentres están mergullados. Igualmente, algunhas familias usan as antenas durante o apareamento e unhas poucas especies úsanas para a defensa. No cerambícido Onychocerus albitarsis, as antenas teñen estruturas que inxectan veleno que utilizan para a defensa.[61] As antenas varían moito en forma, ás veces entre sexos, pero xeralmente son similares dentro dunha mesma familia. As antenas poden ter forma de maza (clavadas), de filamento (filiformes), angulosas (xeniculadas), con forma de corda de doas (moniliformes), de peite (pectinadas nun lado ou nos dous: bipectinadas), ou dentadas (serradas). A variación física das antenas é importante para a identificación de moitos grupos de coleópteros. Os Curculionidae teñen antenas con cóbado ou xeniculadas. As antenas plumosas flabeladas son unha forma restrinxida que se encontra nos Rhipiceridae e algunhas outras familias. Os Silphidae teñen antenas capitadas cunha cabeza esférica no extremo. Os Scarabaeidae teñen antenas lameladas cos segmentos terminais estendidos formando estruturas planas unhas enriba doutras. Os Carabidae teñen antenas filiformes. As antenas orixínanse entre o ollo e as mandíbulas e nos Tenebrionidae, as antenas orixínanse en fronte dunha amosega que rompe o perfil xeralmente circular do ollo composto. Están segmentadas e normalmente constan de 11 partes, a primeira parte denomínase escapo e a segunda parte é o pedicelo. Os outros segmentos denomínanse en conxunto flaxelo.[59][62][63]
As pezas bucais dos coléópteros son como as dos saltóns. As mandíbulas parecen longas pinzas na parte frontal dalgúns escaravellos. As mandíbulas son un par de estruturas duras parecidas a dentes que se moven horizontalmente para agarrar, machacar ou cortar o alimento ou os inimigos (ver defensa, máis abaixo). Dous pares de apéndices parecidos a dedos, os palpos labiais e os maxilares, están situados arredor da boca na maioría dos coleópteros, e serven para mover a comida dentro da boca. En moitas especies, as mandíbulas son sexualmente dimórficas, xa que as dos machos están enormemente agrandads en comparación coas das femias nalgunhas especies.[7]
Tórax[editar | editar a fonte]
O tórax está segmentado en dúas partes discernibles, o pro- e o pterotórax. O pterotórax é o resultado da fusión do meso- e metatórax, que están normalmente separados noutras especies de insectos, aínda que flexiblemente articulados co protórax. Vistos desde abaixo, o tórax é a parte na que se orixinan os tres pares de patas e os dous pares de ás. O abdome é toda a parte do corpo posterior ao tórax.[8] Vistos desde arriba, a maioría dos coleópteros parecen constar de tres claras seccións, mais isto é enganoso: na superficie superior dun escaravello, a sección media é unha placa dura chamada pronoto, que é só a parte frontal do tórax, mentres que a parte posterior do tórax está oculta polas ás do animal. Esta segmentación adicional vese xeralmente mellor no abdome.[64]

Patas[editar | editar a fonte]
As patas multisegmentadas rematan en de dous a cinco pequenos segmentos chmados tarsos. Como en moitas outras ordes de insectos, os coleópteros teñen garras, normalmente un par, ao final do último segmento tarsal de cada pata. Aínda que a maioría dos coleópteros usan as súas patas para camiñar, estas poden estar adaptadas a outros usos. Nos escaravellos acuáticos, como os Dytiscidae, Haliplidae e moitas especies dos Hydrophilidae, as patas, xeralmente o último par, están modificaas para a natación, normalmente con ringleiras de pelos longos. Os escaravellos mergulladores machos teñen copas suctoras nas súas patas anteriores para agarrar as femias.[65] Outros coleópteros teñen patas fosoriais ensanchadas e a miúdo con espiñas para cavar. As especies con esas adaptacións encóntranse nos escarabeidos, carábidos e histéridos. As patas posteriores dalgúns coleópteros, como os Alticini (uns crisomélidos) e os gurgullos saltadores (uns curculiónidos), teñen os fémures agrandados, o que lles facilita choutar.[66]
Ás[editar | editar a fonte]
.jpg)
As ás anteriores non as usan para voar, senón que forman os élitros duros, que cobren a parte posterior do corpo e protexen as ás posteriores. Os élitros son xeralmente estruturas duras cobertoras que deben ser elevadas para permitir que as ás posteriores se movan para poder voar.[67] Porén, nos Cantharidae, os élitros son moles.[68] Outros escaravellos con ás moles son os Lycidae da especie Calopteron discrepans, que teñen ás fráxiles que rompen doadamente para liberar substancias químicas para a defensa.[69]
As ás de voo dos coleópteros están atravesadas por veas e son pregadas unha vez que o animal aterra, a miúdo préganse ao longo desas veas, e quedan gardadas baixo os élitros. Un pregamento (jugum) da membrana na base de cada á é moi característico.[67] Algúns coleópteros perderon a capacidade de voar. Entre estes están algúns carábidos e algúns gurgullos (curculiónidos), e as especies de desertos e covas doutras familias. Moitas teñen os dous élitros fusionados, formando un escudo sólido sobre o abome. Nunhas poucas familias desaparceron tanto a capacidade de voar coma os élitros, como ocorre nos bioluminescentes Phengodidae, nos que as femias lembran larvas durante toda a súa vida.[70] A presenza de élitros e ás non sempre indica que o animal realmente adoite voar. Por exemplo, os Chrysolina graminis (crisomélidos) camiñan dun hábitat a outro malia que fisicamente teñen a capacidade de voar.[71]
Abdome[editar | editar a fonte]
O abdome é a sección que está detrás do metatórax, formada por unha serie de aneis, cada un cun burato respiratorio, chamado espiráculo, compoñendo tres escleritos segmentadoss: o tergo, a pleura e o esterno. O tergo en case todas as especies é membranoso ou normalmente brando e agochado baixo as ás voadoras e élitros cando o animal non voa. As pleuras son xeralmente pequenas ou ocultas nalgunhas especies, e cada unha ten un só espiráculo. O esterno é a parte máis amplamente visible, e pode estar máis ou menos esclerotizado. O abdome non ten apéndices, pero algúns (por exemplo, os Mordellidae) teñen lobos esternais articulados.[72]
Anatomía e fisioloxía[editar | editar a fonte]

Sistema dixestivo[editar | editar a fonte]
O sistema dixestivo dos coleópteros está principalmente adaptado a unha dieta herbívora. A dixestión ten lugar principalmente na arte anterior do intestino medio, aínda que en grupos predadores como os Carabidae, a maioría da dixestión ten lugar no boche por medio dos encimas do intestino medio. Nos Elateridae, as larvas aliméntanse de líquidos que dixiren extraoralmente a comida segregando encimas.[8] A canle alimentaria consiste basicamente nunha farinxe estreita e curta, unha expansión ampla ou boche, e unha escasamente desenvolvida moella. Despois vén o intestino medio, que varía en dimensións entre especies, cun cego de grandes dimensións, e o intestino posterior, de lonxitude variada. Hai normalmente de catro a seis túbulos de Malpighio.[7]
Sistema nervioso[editar | editar a fonte]
O sistema nervioso nos coleópteros pode ser de todos os tipos atopados nos insectos, e varía segundo a especie, desde tres ganglios torácicos e sete ou oito ganglios abdominais, que poden distinguirse do tipo no que todos os ganglios torácicos e abdominais están fusionados para formar unha estrutura composta.[8]
Sistema respiratorio[editar | editar a fonte]

Como a maioría dos insectos, os coleópteros inhalan aire para obter oxíxeno, e exhalan dióxido de carbono a través do sistema traqueal. O aire entra no corpo a través dos espiráculos, e circula por un sistema de traqueas e traqueolas, desde onde difunden os gases ás células.[8]
Os escaravellos mergulladores, como os Dytiscidae, transportan unha burbulla de aire con eles cando se mergullan. Dita burbulla pode quedar retida baixo os élitros ou contra o corpo por pelos hodrofóbicos especializados. A burbulla cobre polo menos algúns dos espiráculols, permitindo que o aire entre nas traqueas.[8] A función da burbulla non só contén unha provisión de aire, senón que tamén actúa como unha branquia física (plastrón). O aire que atrapa está en contacto con auga oxixenada, así que o consumo de oxixeno que fai o animal fai diminuír o oxíxeno da burbulla, e dese modo pode difundir máis oxíxeno desde a auga para reencher a burbulla.[73] O dióxido de carbono é máis soluble en auga que o oxíxeno ou o nitróxeno, polo que difunde con facilidade máis rapidamente cara ao exterior que cara ao interior. O nitróxeno é o gas máis abundante na burbulla e o menos soluble, polo que constitúe un compoñente relativamente estático da burbulla e actúa como un medio estable para que os gases respiratorios se acumulen e atravesen. Unhas visitas ocasionais á superficie do animal son suficientes para que o escaravello restableza a constitución da burbulla.[74]
Sistema circulatorio[editar | editar a fonte]
Como outros insectos, os coleópteros teñen sistemas circulatorios abertos, nos que circula hemolinfa en vez de sangue. Un corazón tubular segmentado está adherido á parede dorsal do hemocele. Ten pares de entradas ou ostiolos a intervalos ao longo da súa lonxitude, e fai circular a hemolinfa desde a principal cavidade do hemocele e fóra a través da cavidade anterior na cabeza.[75]
Órganos especializados[editar | editar a fonte]
Diferentes glándulas están especializadas en producir diversas feromonas, que serven para atraer parellas. As feromonas de certas especies de Rutelinae son producidas polas células epiteliais que tapizan a superficie interna dos segmentos abdominais apicais; as feromonas baseadas en aminoácidos dos Melolonthinae prodúcense en glándulas eversibles do ápice abdominal. Outras especies producen diferentes tipos de feromonas. Os Dermestidae producen ésteres, e algunhas especies dos Elateridae producen aldehidos derivados de ácidos graxos e acetatos.[8] Para atraer as parellas, os vagalumes da familia Lampyridae usan células do corpo graxo modificadas con superficies transparentes que conteñen cristais de ácido úrico reflectantes para reflectir a luz que producen por bioluminescencia. A produción de luz é moi eficiente, e débese á oxidación da luciferina catalizada por encimas luciferases en presenza de adenosín trifosfato (ATP) e oxíxeno, producindo oxiluciferina, dióxido de carbono e luz.[8]
En dúas familias hai órganos timpánicos ou da audición, que consisten nunha membrana (o tímpano) estendida a través dun marco que ten detrás un saco de aire e neuronas sensoriais asociadas.[76] Varias especies do xénero Cicindela (Carabidae) teñen órganos auditivos nas superficies dorsais dos seus primeiros segmentos abdominais debaixo das ás; dúas tribos dos Dynastinae (da familia Scarabaeidae) teñen órganos auditivos xusto debaixo dos seus escudos pronotais ou membranas do pescozo. En ambas as familias eses órganos son sensibles a frecuencias ultrasónicas, cunha forte evidencia que indica que funcionan para detectar a presenza de morcegos pola súa ecolocación ultrasónica.[8]
Reprodución e desenvolvemento[editar | editar a fonte]
Os coleópteros son membros da superorde Endopterygota, e en consecuencia a maioría deles sofren unha metamorfose completa. A forma típica de metamorfose nos coleópteros pasa a través de catro estadios principais: ovo, larva, pupa e imago ou adulto. Nalgunhas especies as pupas poden estar encerradas nun casulo construído pola larva cara ao final do seu último ínstar. Algúns escaravellos, como os típicos membros das familias Meloidae e Ripiphoridae, van un pouco máis alá, experimentando unha hipermetamorfose, na cal o primeiro ínstar ten a forma dun triungulino.[77]
Apareamento[editar | editar a fonte]

Algúns coleópteros teñen un intricado comportamento de apareamento. A comunicación por feromonas adoita ser importante para localizar unha parella. Diferentes especies usan diferentes feromonas. Escaravellos como os Rutelinae usan as feromonas derivadas da síntese de ácidos graxos, mentres que outros, como os Melolonthinae, usan as derivadas de aminoácidos e terpenoides. Outro xeito mediante o cal os escaravellos encontran parella é o dos vagalumes Lampyridae, que son bioluminescentes, con órganos abdominais que producen luz. Os machos e femias establecen un complexo diálogo antes de aparearse; cada especie ten unha combinación única de padróns de voo, duración, composición, e intensidade da luz producida.[8]
Antes de apareárense, machos e femias poden estridular, ou facer vibrar os obxectos sobre os que están. Nos Meloidae, o macho gabea polo dorso da femia e acariña coas súas antenas a cabeza, palpos e antenas dela. En Eupompha, o macho dirixe as súas antenas ao longo do seu vértice lonxitudinal. Pode que non se apareen en toda a vida se non realizan o ritual precopulatorio.[8] O comportamento de apareamento pode ser diferente entre poboacións dispersas da mesma especie. Por exemplo, o apareamento dunha poboación de Rusia do escaravello Chrysolina graminis é precedido por un elaborado ritual no que o macho dá pequenos toques coas súas antenas sobre os ollos, o pronoto e antenas da femia, o cal non é evidente na poboación desta especie no Reino Unido.[78]
A competición pode desempeñar un papel nos rituais de apreamento de coleópteros como os Nicrophorus, no que os insectos loitan para determinar que pode aparearse. Moitos coleópteros machos son territoriais e defenden ferozmente os territorios dos machos intrusos. En tales especies, o macho a miúdo ten cornos na cabeza ou tórax, o que fai que a lonxitude do seu corpo sexa maior que o da femia. A copulación é xeralmente rápida, pero nalgúns casos dura varias horas. Durante a copulación, os espermatozoides son transferidos á femia para fertilizar os ovos.[7]
Ciclo vital[editar | editar a fonte]

Ovo[editar | editar a fonte]
Esencialmente todos os coleópteros poñen ovos, aínda que algúns Aleocharinae e algúns Chrysomelinae mirmecófilos que viven en montañas ou no subártico pode dicirse que son ovovivíparos, xa que poñen ovos que eclosionan case inmediatamente. Os ovos dos coleópteros xeralmente teñen superficies lisas e son brandos, aínda que os Cupedidae teñen ovos duros. Os ovos varían amplamente segundo a especie: adoitan ser pequenos en especies con moitos ínstares (estadios larvarios), e naqueles que poñen un gran número de ovos. Unha femia pode poñer desde ducias a varios miles de ovos durante a súa vida, dependendo do grao de coidados parentais. Estes coidados van desde as especies que simplemente poñen os ovos baixo unha folla aos maiores coidados parentais que prodigan os escarabeidos, que albergan, alimentan e protexen as crías. Os Attelabidae enroscan follas formando un rolo e poñen os seus ovos dentro do rolo para que estean protexidos.[8][79]
Larva[editar | editar a fonte]
A larva é normalmente o principal estadio de alimentación do ciclo vital dos coleópteros. As larvas adoitan alimentarse vorazmente unha vez que emerxen dos ovos. Algunhas aliméntanse externamente sobre as plantas, como as de certos crisomélidos, mentres que outras se alimentan desde o interior das súas fontes de alimento. Exemplos dos que se alimentan no interior son a maioría dos bupréstidos e os cerambícidos. As larvas de moitas familias de coleópteros son predadores de adultos (carábidos, coccinélidos, estafilínidos). O período larvario varía entre as especies, mais pode ser de varios anos. As larvas dos derméstidos experimentan certo grao de reversión do desenvolvemento cando están pasando fame, e posteriormente crecen outra vez ao nivel de madurez que alcanzaran previamente. O ciclo pode ser repetido moitas veces.[80] A morfoloxía larvaria é moi variada entre as distintas especies; poden ter cabezas ben desenvolvidas e esclerotizadas, segmentos torácicos e abdominais distinguibles (usualmente o décimo, aínda que ás veces o oitavo ou noveno).[7]
.jpg)
As larvas de escaravellos poden diferenciarse doutras larvas de insectos polas súas cabezas endurecidas e a miúdo escurecidas, a presenza de pezas bucais mastigadoras e espiráculos ao longo dos laterais dos seus corpos. Igual que os coleópteros adultos, as larvas teñen aparencias variadas, particularmente se comparamos as de distintas familias. Entre os coleópteros con larvas algo aplanadas e moi móbiles están os carábidos e os estafilínidos; as súas larvas descríbense como campodeiformes. Algunhas larvas de coleópteros lembran vermes endurecidos con cabezsas con aspecto de cápsulas escuras e patas diminutas. Hai larvas elateriformes, e encóntranse nos elatéridos e tenebriónidos. Os escaravellos Scarabaeoidea teñen larvas grosas e curtas descritas como escarabaeiformes.[81]
Todas as larvas de coleópteros pasan por varios ínstares, que son os estadios de desenvolvemento entre cada muda. En moitas especies, as larvas simplemente incrementan o seu tamaño con cada ínstar sucesivo a medida que consomen máis alimento. Nalgúns casos, non obstante, ocorren cambios máis drásticos. Entre certas familias ou xéneros, especialmente os que mostran estilos de vida parasitos, o primeiro ínstar (o planidio) é moi móbil para poder buscar un hóspede, mentres que os seguintes son moi sedentarios e permanecen sobre ou dentro do seu hóspede. Isto coñécese como hipermetamorfose; ocorre nos Meloidae, Micromalthidae e Ripiphoridae.[82] O escaravello Epicauta vittata (Meloidae), por exemplo, ten tres estadios larvarios. O seu primeiro estadio, o triungulino, ten patas longas para ir en busca de ovos de saltóns, que come. Despois de comer durante unha semana, muda ao segundo estadio, chamado o estadio caraboide, que lembra a larva dos Carabidae. Unha semana despois muda e adopta a aparencia dunha larva de Scarabaeidae, chamada estadio escarabaeidoide. O seu penúltimo estadio larvario é a pseudopupa ou larva coarcada, que inverna e fai a pupación ata a seguinte primavera.[83]
O período larvario pode variar amplamente. O estafilínido comedor de fungos Phanerota fasciata experimenta tres mudas en 3,2 días a temperatura moderada, mentres que Anisotoma sp. (Leiodidae) completa o seu estadio larvario no corpo frutífero de mofos mucosos en 2 días e posiblemente representa o crecemento máis rápido entre os coleópteros. O escaravello derméstido Trogoderma inclusum pode permanecer nun estado larvario ampliado baixo condicións desfavorables, incluso reducindo o seu tamaño entre mudas. Informouse dunha larva que sobreviviu 3,5 anos nun recipiente pechado.[8]
Pupa e adulto[editar | editar a fonte]

Como todos os endopterigotos, as larvas dos coleópteros pupan, e destas pupas emerxen adultos sexualmente maduros e completamente formados ou imagos. As pupas nunca teñen mandíbulas (son adecticosas). Na maioría das pupas, os apéndices están libres e non unidos en toda a súa lonxitude ao corpo e denomínanse exaratas; nuns poucos escaravellos (Staphylinidae, Ptiliidae etc.) os apéndices están fusionados ao corpo (denominados pupas obtectas).[7]
Os adultos teñen duracións da vida extremadamente variables, de semanas a anos, dependendo das especies.[7][46] Algúns escaravellos perforadores da madeira poden ter ciclos de vida extremadamente longos. Crese que cando os mobles ou as madeiras da casa están infestados por larvas destes escaravellos, a madeira xa contiña as larvas cando foi inicialmente serrada. Informouse que dunha caixa de madeira de bidueiro de 40 anos de antigüidade liberouse un adulto de Eburia quadrigeminata (un Cerambycidae), mentres que se documentou que adultos de Buprestis aurulenta e outros Buprestidae emerxeron 51 anos despois da fabricación de obxectos de madeira que contiñan as súas larvas.[84]
Comportamento[editar | editar a fonte]
Locomoción[editar | editar a fonte]

Os élitros permiten que os coleópteros poidan voar e tamén moverse en espazos estreitos, o que fan pregando as súas delicadas ás voadoras baixo os duros élitros cando non están voando, e despregando as ás voadoras só xusto antes de engalar. O pregamento e despregamento das ás é realizado pola acción de músculos unidos á base das ás; con tal de que se manteña a tensión nas veas radial e cubital, as ás permanecen estiradas. Nalgunhas especies que voan polo día (por exemplo, os Buprestidae ou os Scarabaeidae), o voo non precisa unha gran sustentación por parte dos élitros, ao ter as ás metatorácicas estendidas baixo as marxes laterais dos élitros.[8] A altitude en voo alcanzada polos coleópteros varía. Un estudo que investigou isto nas especies de maruxiñas Coccinella septempunctata e Harmonia axyridis usando o radar encontrou que, mentres que maioría dos que voaban sobre unha determinada localidade facíano a 150–195 m sobre o chan, algunhas alcanzaban altitudes duns 1100 m.[85]
Moitos estafilínidos reduciron notablemente os seus élitros e, aínda que teñen a capacidade de voar, o máis normal é que se movan sobre o chan: os seus corpos moles e fortes músculos abdominais fan que sexan moi flexibles e que doadamente poidan escapar meténdose en pequenas gretas.[86]
Os escaravellos acuáticos usan varias técnicas para reter aire baixo a superficie da auga. Os escaravellos mergulladores (Dytiscidae) reteñen o aire entre o abdome e os élitros cando están mergullados. Os Hydrophilidae teñen pelos na súa superficie inferior que reteñen unha capa de aire pegada aos seus corpos. Os escaravellos acuáticos adultos que se arrastran usan tanto os élitros coma as coxas das patas posteriores (segmentos basais das patas) para a retención de aire, mentres que os Gyrinidae simplemente transportan con eles baixo a auga unha burbulla de aire cando se mergullan.[87]
Comunicación[editar | editar a fonte]
Os escravellos teñen diversos modos de comunicarse, incluíndo o uso de feromonas. O escaravello do piñeiro emite unha feromona para atraer a outros escaravellos a unha árbore. A masa de escaravellos pode superar as defensas químicas dunha árbore. Unha vexz que as defensas da árbore se esgotan, os escaravellos emiten unha feromona antiagregación. A especie pode estridular para comunicarse.[88]
Coidados parentais[editar | editar a fonte]

Unhas poucas especies de coleópteros prodigan coidados parentais ás crías, quizais para protexelas das condicións adversas e os predadores.[8] O estafilínido Bledius spectabilis viven en marismas salinas, polo que os ovos e larvas están en perigo pola subida da marea. A nai vixía os ovos e larvas, escavando para preservalos do asolagamento e a asfixia, e protéxeos do escaravello carábido Dicheirotrichus gustavi e da avespa parasitoide Barycnemis blediator, que mata un 15% das larvas.[89]
Os escaravellos enterradores son pais moi atentos e participan nun coidado cooperativo e na alimentación da súa descendencia. Ambos os proxenitores traballan para enterrar pequenas carcasas de animais para que sirvan como fonte de alimento ás crías e contrúen unha cámara de cría arredor delas. Os pais preparan as carcasas e protéxenas dos competidores e dunha descomposición temperá. Despois de que eclosionan os ovos, os parentes manteñen as larvas limpas de fungos e bacterias e axudan á alimentación das larvas regurxitando comida para elas.[90]
Algúns escaravellos peloteiros proporcionan coidados parentais, recollendo excrementos de herbívoros e poñendo ovos nesa fonte de alimento. Algunhas especies non os abandonan nesta fase, senón que permanecen alí para salvagardar a súa descendencia.[91]
Non obstante, a maioría das especies de escaravellos non presentan comportamentos de coidados parentais unha vez que depositaron os ovos.[92]
Eusocialidade[editar | editar a fonte]
A eusocialidade implica coidados cooperativos na crianza (incluíndo o coidado das crías por parte doutros individuos distintos dos proxenitores), o solapamento das xeracións nunha colonia de adultos e a división do traballo entre os grupos de individuos non reprodutores e reprodutores.[93] Hai poucos organismos á parte dos himenópteros que mostren este comportamento; e o único coleóptero que fai isto é o gurgullo Austroplatypus incompertus.[94] Esta especie australiana vive en redes de túneles horizontais na cerna do tronco dos Eucalyptus. É unha das máis de 300 especies de escaravellos da ambrosía (uns gurgullos) perforadores da madeira, que distribúen as esporas dos fungos da ambrosía.[95] Os fungos crecen nos túneles dos coleópteros, proporcionando alimento para os escaravellos e as súas larvas; a descendencia feminina permanece nos túneles e mantén o crecemento dos fungos, e probablemente nunca se reproduce. Só hai unha femia reprodutora.[94][95] Os coidados cooperativos ás crías tamén se encontran nos coleópteros Passalidae, nos que as larvas se alimentan de feces semidixeridas dos adultos.[96]
Alimentacion[editar | editar a fonte]
_on_Ipomoea_carnea_W_IMG_0593.jpg)
Os escaravellos poden acceder a unha gran diversidade de fontes de alimento dispoñibles nos seus moitos hábitats. Algúns son omnívoros e comen plantas e animais. Outros teñen unha dieta moi especializada. Moitas especies de crisomélidos, cerambícidos e gurgullos son moi específicos da súa planta hóspede, alimentándose só dunha especie de planta. Os carábidos e estafilínidos, entre outros, son principalmente carnívoros e capturan e comen moitos outros artrópodos e pequenas presas, como miñocas e caracois. Aínda que a maioría dos escaravellos predadores son xeneralistas, unhas poucas especies consomen presas máis específicas ou teñen preferencias.[97]
A materia orgánica en descomposición é unha dieta primaria para moitas especies. Esta pode ser desde excrementos, que consomen as especies coprófagas (como os escaravellos peloteiros da familia Scarabaeidae), ata animais mortos, que comen as especies necrófagas (Silphidae). Algúns escaravellos que se poden encontrar nos excrementos e na prea son de feito predadores. Entre estes están membros dos Histeridae e Silphidae, que se encontran alí para depredar nas larvas dos insectos coprófagos e necrófagos.[98] Moitos coleópteros aliméntanse de codias de árbores, algúns aliméntanse de madeira e outros de fungos que crecen na madeira ou follas caídas. Algúns coleópteros teñen estruturas especiais chamadas micanxios, para o transporte de esporas fúnxicas.[99]
Ecoloxía[editar | editar a fonte]

Adaptacións antidepredadores[editar | editar a fonte]
Os coleópteros, tanto adultos coma larvas, son as presas de moitos animais predadores, como mamíferos, desde morcegos a roedores, e tamén de aves, lagartos, anfibios, peixes, libélulas, asílidos, redúvidos, formigas, outros coleópteros, e arañas.[100][101] Os coleópteros usan diversas adaptacións antipredador para defenderse. Entre estas está a camuflaxe e o mimetismo contra os predadores que os cazan pola vista, a toxicidade, e o comportamento defensivo.
Camuflaxe[editar | editar a fonte]
- Véxase tamén: Camuflaxe.
A camuflaxe é algo común e moi estendido entre os coleópteros, especialmente nos que se alimentan da madeira e vexetación, como os crisomélidos (que adoitan ser verdes) e os gurgullos. Nalgunhas especies, a feitura do corpo ou pelos e escamas de varias cores fan que os escaravellos, como por exemplo o gurgullo Heilipus apiatus, lembren excrementos de ave ou outros obxectos incomestibles.[100] Moitos coleópteros viven en ambientes areosos e imitan a coloración do substrato.[102]
Mimetismo e aposematismo[editar | editar a fonte]

- Véxase tamén: Mimetismo e Aposematismo.
Algúns cerambícidos son imitadores batesianos das avespas. Os coleópteros poden combinar a coloración co mimetismo de comportamento, actuando como as avespas ás que xa se parecen fisicamente. Moitos outros coleópteros, como as xoaniñas, meloidos e lícidos segregan substancias de moi mal sabor ou tóxicas o que os fai incomestibles ou velenosos e adoitan ser aposemáticos, xa que teñen unha rechamante coloración que advirte os depredadores; moitos escaravellos e outros insectos imitan á súa vez estas especies quimicamente protexidas.[103]
.jpg)

A defensa química é importante nalgunhas especies, e xeralmente a advirten por medio de cores aposemáticos rechamantes. Algúns Tenebrionidae adoptan posturas para liberar compostos químicos noxentos para advertir os depredadores. As defensas químicas poden servir para outras cousas ademais de para protexerse dos vertebrados, como a protección contra diversos microbios. Algunhas especies secuestran compostos químicos das plantas das que se alimentan, incorporándoos ás súas propias defensas.[102]
Outras especies teñen gándulas especiais para producir compostos químicos disuasorios. As glándulas defensivas dos carábidos producen diversos hidrocarburos, aldehidos, fenois, quinonas, ésteres e ácidos liberados desde unha abertura situada ao final do abdome. Carábidos africanos como por exemplo, Anthia e Thermophilum (Thermophilum é ás veces incluído dentro de Anthia) empregan a mesma substancia química que as formigas: o ácido fórmico.[103] Os escaravellos bombardeiros teñen glándulas pixidiais ben desenvolvidas que baleiran desde os lados das membranas intersegmentarias entre os segmentos abdominais sétimo e oitavo. A glándula está formada por dúas cámaras de depósito, unha contén hidroquinonas e peróxido de hidróxeno, e a outra contén peróxido de hidróxeno e o enzima catalase. Estes compostos químicos mestúranse e o resultado é unha exección explosiva, que chega a temperaturas duns 100 °C, na que se degrada a hidroquinona a hidróxeno, oxíxeno e quinona. O oxíxeno propulsa o fedorento spray químico como un chorro que pode dirixirse apuntando con precisión aos predadores.[8]
Outras defensas[editar | editar a fonte]
Os escravellos grandes que viven no chan, como os carábidos, o escaravello rinoceronte e os cerambícidos deféndense usando as súas fortes mandíbulas ou espiñas fortemente esclerotizadas ou cornos para disuadiren ou loitaren contra os predadores.[102] Moitas especies de gurgullos que se alimentan na parte exposta de follas de plantas reaccionan ao ataque empregando un reflexo de tirarse da planta ao chan. Algúns combinan isto coa tanatose, na cal retraen os seus apéndices e "fanse os mortos".[104] Os elatéridos poden catapultarse de repente para escaparen da zona de perigo ao liberaren a enerxía almacenada por un mecanismo de clic, que consta dunha robusta espiña situada no prosterno que se engancha nunha fenda do mesosterno.[100]
Parasitismo[editar | editar a fonte]
Unhas poucas especies de coleópteros son ectoparasitas de mamíferos. Unha delas é Platypsyllus castoris, que parasita castores. Este escaravello vive como parasito tanto na etapa de larva coma na de adulto, alimentándose do tecido epidérmico do castor e posiblemente das secrecións da pel e dos exsudados das feridas. Están moi aplanados dorsoventralmente, o que é unha adaptación para deslizarse entre os pelos dos castores. Non teñen ás nin ollos, como ocorre con moitos ectoparasitos.[105] Outros son cleptoparasitos doutros invertebrados, como o escaravello das colmeas Aethina tumida, que infesta as colmeas de abellas do mel,[106] mentres que outras especies son inquilinas parasitas ou comensais en formigueiros.[107] Un pequeno grupo de escaravellos son principalmente parasitoides doutros insectos, alimentándose e finalmente matando os seus hóspedes.[108]
.jpg)
Polinización[editar | editar a fonte]
As flores polinizadas por coleópteros son xeralmente grandes, verdosas ou abrancazadas e pouco perfumadas. Os aromas poden ser picantes, frutais ou similares ao da materia orgánica en descomposición. Os coleópteros probablemente foron os primeiros insectos que polinizaron flores. A maioría das flores polinizadas por escaravellos son aplanadas ou discoidais, o pole doadamente accesible, aínda que poden ter trampas para a polinización para manter ao escaravellos alí durante máis tempo. Os ovarios da planta están xeralmente protexidos das pezas bucais mordedoras dos seus polinizadores. As familias de coleópteros que polinizan habitualmente flores son as dos Buprestidae, Cantharidae, Carambycidae, Cleridae, Dermestidae, Lycidae, Melyridae, Mordellidae, Nitidulidae e Scarabeidae.[109] Os escaravellos poden ser especialmente importantes nalgunhas partes do mundo como as áreas semiáridas do sur de África e sur de California[110] e as praderías de montaña de KwaZulu-Natal en Suráfrica.[111]
Mutualismo[editar | editar a fonte]

2: A larva aliméntase dos fungos, que dixiren a madeira, eliminando toxinas para mutuo beneficio.
3: Larva pupando.
O mutualismo dáse nalgúns coleópteros, como nos chamados escaravellos da ambrosía (uns gurgullos), que se asocian con fungos para dixerir a madeira de árbores mortas. Os coleópteros escavan túneles nas árbores mortas nas cales cultivan xardíns de fungos, a súa única fonte de nutrición. Despois de aterraren nunha árbore axeitada, un destes coleópteros escava un túnel no cal libera esporas do seu simbionte fúnxico que transporta. O fungo penetra no tecido xlemático da planta, dixíreo, e concentra os nutrientes sobre ou preto da superficie da galería que fixo o animal, así que os gurgullos e o fungo poden beneficiarse ambos. Os gurgullos non poden comer a madeira directamente debido ás toxinas que ten e utilizan as súas relacións cos fungos para axudarlles a superar as defensas da súa árbore morta e proporcionarlle nutición ás súas larvas.[112] Esta relación mutualista entre o coleóptero e o fungo quimicamente mediada por un peróxido poliinsaturado producido por bacterias[113] orixinouse por coevolución.[112][114]
Tolerancia a ambientes extremos[editar | editar a fonte]

Un 90% das especies de coleópteros entran nun período de diapausa adulta, unha fase de repouso cun metabolismo reducido para afrontar as condicións ambientais desfavorables. Esta diapausa dos adultos é a forma máis común de diapausa nos coleópteros. Para resistir o período sen alimentos (que a miúdo pode durar varios meses) os adultos prepáranse acumulando reservas de lípidos, glicóxeno, proteínas e outras substancias necesarias para superar futuros cambios perigosos nas condicións ambientais. Esta diapausa é inducida por sinais que anuncian a chegada da estación desfavorable; normalmente a pista é un cambio no fotoperíodo. Os días curtos (en diminución) serven como sinal de que se aproxima o inverno e inducen a diapausa invernal (hibernación).[115] Un estudo da hibernación no escaravello ártico Pterostichus brevicornis mostrou que os niveis de graxa corporal dos adultos eran os máximos en outono e tiñan entón a canle alimentaria chea de comida, pero estaba baleira a finais de xaneiro. Esta perda da graxa corporal era un proceso gradual, que ocorría en combinación cunha deshidratación.[116]
Todos os insectos son poiquilotermos,[117] polo que a capacidade dunhas poucas especies de escaravellos de viviren en ambientes extremos depende da súa resistencia a temperaturas especialmente altas ou baixas. O escaravello Pityogenes chalcographus pode sobrevir a −39 °C mentres pasa o inverno baixo a codia dunha árbore;[118] o escaravello de Alasca Cucujus clavipes puniceus pode soportar os −58 °C, e as súas larvas poden sobrevivir a −100 °C.[119] A estas baixas temperaturas, a formación de cristais de xeo nos fluídos internos é a maior ameaza para a supervivencia dos escaravellos, pero isto impídese pola produción de proteínas anticonxelantes que impiden que as moléculas de auga se agrupen. As baixas temperaturas experimentadas por Cucujus clavipes poden ser soportadas pola deliberada deshidratación xunto coas mencionadas proteínas anticonxelantes. A deshidratación multiplica a concentración destes anticonxelantes.[120] A hemolinfa do escaravello Tenebrio molitor contén varias proteínas anticonxelantes.[121] O escaravello de Alasca Upis ceramboides pode sobrevivir a −60 °C: os seus crioprotectores son xilomananos, moléculas que constan deun azucre unido a un ácido graxo,[122] e o azucre-alcohol treitol.[123]
Inversamente, os escaravellos que viven nos desertos están adaptados a tolerar altas temperaturas. Por exemplo, o escaravello tenebriónido Onymacris rugatipennis pode soportar os 50 °C.[124] Os Cicindelinae en áreas areosas quentes son a miúdo de tons brancos (por exemplo, Habroscelimorpha dorsalis), para así reflectir máis calor. Estes escaravellos tamén presentan adaptacións de comportamento para tolerar a calor: poden erguerse sobre os seus tarsos para manter o seu corpo a máis afastado posible do chan, procuran sitios de sombra, e volven a cabeza cara ao sol para que só a parte frontal da súa cabeza quede directamente exposta.[125]
O escaravello do deserto namibio Stenocara gracilipes pode recoller auga da néboa, xa que os seus élitros teñen unha superficie con textura na que se combinan protuberancias hidrófilas (que captan auga) e depresións cerosas hidrófobas. O escaravello exponse á brisa de mañanciña, elevando o seu abdome; condénsanse pingas de auga sobre os seus élitros que flúen ao longo de rugosidades cara á boca. Adaptacións similares encóntranse noutros escaravellos do deserto de Namibia como Onymacris unguicularis.[126]
Algúns escaravellos terrestres que explotan os hábitats costeiros e de chaira inundable teñen adaptacións fisiolóxicas para sobrevivr ás inundacións. No caso dunha inundación, os adultos poden ter mobilidade dabondo como para afastarse rapidamente da zona que se está a asolagar, pero as larvas e pupas xeralmente non. Os adultos de Cicindela togata non poden sobrevivir á inmersión na auga, mais as larvas poden sobrevivir durante un período prolongado de ata seis días de anoxia durante as inundacións. A tolerancia á anoxia nas larvas pode deberse a que cambian a un metabolismo anaerobio ou a que reducen a taxa metabólica.[127] A tolerancia á anoxia no carábido adulto Pelophila borealis foi comprobada en condicións de laboratorio e atopouse que podía sobrevivir a un período continuado de 127 días nunha atmosfera de 99,9% de nitróxeno a 0 °C.[128]
Migración[editar | editar a fonte]
Moitas especies de coleópteros realizan movementos en masa anuais, que se denominan migracións. Entre estas están o Meligethes aeneus[129] e moitas especies de coccinélidos.[130] Estes movementos en masa poden tamén ser oportunistas, para a procura de alimento, en vez de estacionais. Un estudo de 2008 dunha praga especialmente grande de escaravello do piñeiro (Dendroctonus ponderosae) na Columbia Británica encontrou que os escaravellos podían voar de 30 a 110 km diarios en densidades de ata 18 600 escaravellos por hectárea.[131]
Relacións cos humanos[editar | editar a fonte]
Nas culturas antigas[editar | editar a fonte]
- Artigo principal: Escarabeo.

Varias especies de escaravellos peloteiros, especialmente o escaravello sagrado, Scarabaeus sacer, eran reverenciados no Antigo Exipto.[132][133] A imaxe xeroglífica do escaravello crese que tiña unha importancia existencial, de ficción, ou ontolóxica.[134] Atopáronse imaxes de escarabeos en ósos, marfil, pedras, cerámica e metais preciosos desde a sesta dinastía e ata o período de dominio romano. O escarabeo era de primordial importancia no culto funerario do Antigo Exipto.[135] O escarabeo estaba vinculado a Khepri, o deus do sol nacente, debido á suposta semellanza entre o rodar unha bóla de excrementos que fai o escaravello e o rodar do sol que fai o deus.[132] Algúns dos veciños da antiga civilización exipcia adoptaron o motivo do escarabeo para selos de varios tipos. Os mellor coñecidos son os selos LMLK xudeus, nos que oito dos seus 21 debuxos representan escarabeos, que eran usados para estampar impresións en tinallas de almacenamento durante o reinado de Ezequías.[136] Os escaravellos son mencionados como símbolo do sol, igual que no Antigo Exipoto, na obra de Plutarco do século I Moralia.[137] O Papiro máxico grego do século II a.C. ao século V d.C. describe os escarabeos como un ingrediente nun encantamento.[138]
Moitos séculos despois o escarabeo pasou a ter un valor simbólico no cristianismo copto. Como os antigos exipcios considerábano un exemplo da vida eterna, ao supoñeren erradamente que 'resucitaba' da bóla de excrementos que facía, situaban sobre o corazón das momias unha imitación do escaravello. Posteriormente o cristianismo copto asimilou sincreticamente aspectos da relixión precedente, e, ademais de adoptar a cruz ansada, tamén adoptou -como alegoría- o escarabeo, asimilándoo metaforicamente a Xesucristo, que tamén resucitara, ao que chamaban "O Bo Escaravello", por iso nalgúns textos de Agostiño de Hipona e posteriores poden encontrarse expresións como "bonus scarabæus" en referencia a Cristo.
Plinio o Vello trata dos escaravellos na súa Historia Natural,[139] describindo a vacaloura: "Algúns insectos, para a conservación das súas ás, están cubertos cun erusto (élitros) – o escaravello, por exemplo, cuxas ás son peculiarmente finas e fráxiles. A estes insectos a Natureza negoulle ter un aguillón; pero nunha gran clase deles encontramos que teñen cornos de singular lonxitude, con dous dentes nos extremos, formando pinzas, que o animal pecha cando ten a intención de trabar."[140][141] A vacaloura aparece nun mito grego de Nicander lembrado por Antonino Liberalis no cal Cerambus[b] é convertido en escaravello: "Pode verse sobre troncos e ten dentes ganchudos, movendo sempre as súas mandíbulas á vez. É negro, longo e ten ás duras como o gran escaravello peloteiro".[142] A historia conclúe co comentario de que os escaravellos eran usados como xoguetes polos rapaces, e que lle quitaban a cabeza e usábana como un colgante.[141][143]
Etnografía galega[editar | editar a fonte]
Ditos sobre o escaravello na cultura galega son:[144].
- Ao escaravello, os seus fillos parécenlle ouro fino.
- O que fixer de min escaravello, non farei del espello.
Como pragas[editar | editar a fonte]

Un 75% das especies de escaravellos son fitófagas tanto en estado larvario coma adulto. Moitos comen plantas economicamente importantes e produtos vexetais almacenados, como árbores, cereais, tabaco e froitos secos.[7] Algúns, como o gurgullo do algodoeiro (Anthonomus grandis), que se alimenta de xemas das flores do algodoeiro, poden causar danos moi graves á agricultura. Este gurgullo cruzou o Río Grande por Texas, e entrou nos Estados Unidos desde México contra 1892,[145] e xa alcanzara o sueste de Alabama en 1915. A mediados da década de 1920, entrara en todas as rexións algodoeiras dos Estados Unidos, viaxando de 60 a 260 km por ano. Hoxe segue sendo a praga máis destrutiva para en Norteamérica. A Universidade do Estado de Mississippi estimou, que desde que este insecto entrou no país supuxo un custo para os produtores de algodón de 13 mil millóns de dólares, e nos tempos máis recentes duns 300 millóns de dólares ao ano.[145]

Unha praga moi importante é o escaravello da pataca, Leptinotarsa decemlineata, cuxas larvas (as máis voraces) e adultos comen as follas da pataca. Os seus hóspedes poden ser tamén outros membros da familia Solanaceae, como o tomate, berenxena, pemento e outros. É nativo de América, pero espallouse por Europa e parte de Asia.[146][147] A praga apareceu en 1859 na rexión de Omaha en Nebrasca. Espallouse rapidamente e en 1874 xa chegara á costa atlántica norteamericana,[146] e en 1877 chegou ao Reino Unido, onde houbo desde entón sucesivas erradicacións e repuntas.[148] No resto de Europa o escaravello entrou na base militar americana en Bordeos, Francia durante ou pouco despois da Primeira guerra mundial e desde entón non deixou de espallarse e agora encóntrase en case todo o continente. Na Unión Europea actualmente a praga non está establecida en ningún país membro, pero poden ocorrer infestacións ocasionais.[149][150] No escaravello da pataca evolucionou unha inmunidade aos insecticidas. Diferentes poboacións desta especie desenvolveron resistencia a todas as principais clases de insecticidas.[151] O escaravello da pataca foi avaliado como ferramenta para a guerra entomolóxica durante a Segunda guerra mundial, coa intención de usar as súas larvas para danar as colleitas das nacións inimigas.[152] Alemaña probou o seu programa de utilización como arma deste escaravello en Frankfurt, liberando 54 000 escaravellos.[153]
Coleópteros que causan danos nos olmos son algúns Scolytidae, o crisomélido Xanthogaleruca luteola e o cerambícido asiático Anoplophora glabripennis[154]. Os Scolytidae transmiten o fungo que produce a grafiose cando se desprazan de zonas infectadas a zonas sas. Esta enfermidade devastou os olmos de Europa e Norteamérica.[155]
O escaravello anóbido Xestobium rufovillosum é unha seria praga para as construcións de madeira vellas en Europa. Ataca as madeiras duras como a dos carballos e castiñeiros, sempre por partes onde se produciu unha putrefacción por fungos. A introdución destas pragas nos edificios crese se se produce no momento da construción.[156]
Outros coleópteros que causan pragas son os escaravellos do coqueiro, Brontispa longissima, que se alimenta de follas nacentes, plantas xoves e árbores maduros do coqueiro, causando graves danos económicos en Filipinas.[157] O escaravello do piñeiro (Dendroctonus ponderosae) é unha destrutiva praga dos piñeiros Pinus contorta maduros ou debilitados, e ás veces afecta grandes áreas do Canadá.[158]
Como recursos beneficiosos[editar | editar a fonte]

Os escaravellos poden ser beneficiosos para a economía humana para controlar as poboacións de especies que causan pragas. As larvas e adultos dalgunhas especies de maruxiñas (Coccinellidae) aliméntanse de áfidos que son nocivas pragas. Outras xoaniñas aliméntanse de hemípteros Coccoidea (cochinillas), Aleyrodidae e Pseudococcidae.[159] Se escasean as fontes normais de alimento, poden alimentarse de pequenas eirugas, míridos xoves, ou das secrecións doutros insectos (melada) e néctar.[160] Os carábidos son predatores comúns de moitos insectos que son pragas, incluíndo ovos de moscas, eirugas e elatéridos.[161] Os carábidos poden axudar tamén a controlar malas herbas ao comeren as súas sementes no chan, reducindo a necesidade de usar herbicidas para protexer colleitas.[162] A efectividade dalgunhas especies para reducir certas poboacións de plantas tivo como resultado a introdución deliberada de escaravellos para controlar as malas herbas. Por exemplo, o xénero Zygogramma é nativo de Norteamérica pero foi utilizado para controlar a planta Parthenium hysterophorus na India e a planta Ambrosia artemisiifolia en Rusia.[163][164]
Os escaravellos peloteiros (Scarabidae) foron utilizados con éxito para reducir a poboación de moscas como Musca vetustissima e Haematobia exigua, que son graves pragas para o gando vacún en Australia.[165] Os escaravellos fan que estas moscas non teñan excrementos para reproducirse neles ao faceren bólas con eles e enterralas rapidamente, o que ademais fertiliza o chan, e facilita o ciclo dos nutrientes.[166] O Australian Dung Beetle Project (1965–1985), introduciu especies de escaravello peloteiro en Australia desde Suráfrica e Europa para reducir as poboacións de Musca vetustissima, ao que seguiron sucesivos ensaios desta técnica en Hawai.[165] O Instituto Americano de Ciencias Biolóxicas informou que os escaravellos peloteiros aforraron á industria gandeira estadounidense uns 380 millóns de dólares anualmente ao enterraren as feces do gando vacún.[167]
Os Dermestidae utilízanse a miúdo en taxidermia e na preparación de espécimes científicos, para que limpen de tecidos brandos os ósos.[168] As larvas aliméntanse e eliminan a cartilaxe e outros tecidos brandos.[169][170]
Como alimento[editar | editar a fonte]

Os escaravellos son os insectos máis amplamente consumidos como alimento polos humanos, e hai unhas 344 especies usados como comida, xeralmente no estado larvario.[171] Os Tenebrio molitor (larvas de tenebriónidos) e o escaravello rinoceronte están entre as especies que comunmente se comen.[172]
Como indicadores de biodiversidade[editar | editar a fonte]
Debido á súa especificidade de hábitat, moitas especies de coleópteros foron propostas como indicadores ambientais axeitados, xa que a súa presenza, cantidade ou ausencia proporciona unha mededa da calidade do hábitat. Os escaravellos predadores como os Cicindelidae teñen un uso científico como taxons indicadores para medir os padróns rexionais de biodiversidade. Son adecuados para isto xa que a súa taxonomía é estable; a súa historia vital está ben descrita; son grandes e doados de observar cando se visita un lugar; aparecen por todo o mundo en moitos hábitats, con especies especializadas en determinados hábitats; e a aparición de determinadas especies indica con precisión a presenza doutras especies, tanto de vertebrados coma de invertebrados.[173] Segundo os hábitats, moitos outros grupos como os estafilínidos en hábitats modificados polos humanos, os escaravellos peloteiros nas sabanas[174] e os escaravellos saproxílicos (comedores de madeira) en bosques[175] foron propostos como especies potencialmente indicadoras.[176]
Na arte e adornos[editar | editar a fonte]


Moitos escaravellos teñen élitos fermosos e duradeiros que foron usados como materiais na arte, por exemplo nos adornos con élitros asiáticos.[177] Ás veces foron incorporados como obxectos rituais de importancia relixiosa. Os escaravellos completos, xa sexa como tales ou con cubertas de plástico, convertéronse en obxectos vendidos como souvenirs, desde chaveiros a requintadas xoias caras. En partes de México, os escaravellos do xénero Zopherus convértense en broches vivos ao pegarlles xoias de fantasía e cadeas de ouro, o que é posible grazas á incrible dureza dos élitros e aos hábitos sedentarios deste xénero.[178]
En espectáculos[editar | editar a fonte]
Os escaravellos loitadores utilízanse en partes de Asia como espectáculo e para facer apostas. Este "deporte" explota o comportamento territorial e a competición polo apareamento de certas especies de escaravellos grandes. No distrito de Chiang Mai do norte de Taiandia, captúranse na natureza machos de escaravellos rinoceronte Xylotrupes, que son adestrados para a loita. As femias mantéñense dentro dun tronco para estimular a loita dos machos coas súas feromonas.[179] Estas loitas poden ser competitivas e fanse apostas de diñeiro ou propiedades por quen vai ser o gañador.[180] En Corea do Sur a especie de ditíscidos Cybister tripunctatus utilizase nun xogo parecido á ruleta.[181]
Os escaravellos son ás veces utilizados como instrumentos musicais: o pobo onabasulu de Papúa Nova Guinea historicamene usaba o gurgullo Rhynchophorus ferrugineus como instrumento musical deixando que a cavidade bucal humana servise como cámara de resoancia variable para as vibracións feitas coas ás polo insecto adulto vivo.[180]
Como mascotas[editar | editar a fonte]
Algunhas especies de escaravellos téñense en casa como mascota, como é o caso dos escaravellos acuáticos Dytiscidae, que se poden manter nun tanque de auga doce nas casas.[182]

No Xapón, a práctica de ter como mascotas escaravellos rinoceronte cornudos (Dynastinae) e vacalouras (Lucanidae) é especialmente popular entre os rapaces.[183] Tal é a popularidade disto no Xapón que en 1999 deseñáronse máquinas expendedoras automáticas que dispensan os escaravellos vivos, cada unha das cales pode conter 100 vacalouras.[184][185]
Como obxectos de colección[editar | editar a fonte]
A colección de escaravellos foi moi popular na era vitoriana.[186] O naturalista Alfred Russel Wallace recolleu (pola súa conta) un total de 83 200 escaravellos durante oito anos tal como describiu no seu libro de 1869 The Malay Archipelago (O arquipélago malaio), incluíndo 2 000 especies novas para a ciencia.[187]
Como inspiración para certas tecnoloxías[editar | editar a fonte]
Varias adaptacións dos coleópteros suscitaron o interese dos investigadores en biomimética con posibles aplicacións comerciais. O spray repelente do escaravello bombardeiro inspirou o desenvolvemento da tecnoloxía dun spray de nebulización fina que se considera que ten un baixo impacto de carbono comparado cos sprays de aerosol.[188] O comportamente de recollida de humidade dos escaravellos do deserto do Namib (Stenocara gracilipes) inspirou o deseño dunha botella de auga que se autoenche, que utiliza materiais hidrófilos e hidrófobos para beneficiar á xente que vive en rexións secas sen chuvias regulares.[189]
Os escaravellos vivos foron utilizados como cyborgs. Un proxecto financiado pola Axencia de Proxectos de Investigación Avanzada (ARPA) dos Estados Unidos implantou eléctrodos no escaravello Mecynorhina torquata, por medio dos cales se podía controlalo remotamente cun receptor de radio que o animal levaba ás costas, como proba do concepto para traballos de vixilancia.[190] Unha tecnoloxía similar aplicouse para permitir que un operador humano controle o rumbo en voo libre e o paso ao camiñar de Mecynorhina torquata, así como os xiros graduais e camiñar cara a atrás de Zophobas morio.[191][192][193]
En conservacionismo[editar | editar a fonte]
Como os escaravellos constitúen unha parte tan grande da biodiversidade mundial, a súa conservación é importante, e igualmente, a perda de hábitat e biodiversidade con certeza causa un impacto sobre os coleópteros. Desafortunadamente, con excepcións para grupos como as xoaniñas e vagalumes, os escaravellos teñen en conxunto unha imaxe menos positiva que outros grupos de insectos como, por exemplo, as bolboretas: moitos deles causan pragas agrícolas, e outros causan noxo. Moitas especies de escaravellos teñen hábitats moi específicos e ciclos de vida longos que os fan vulnerables. Algunhas especies están moi ameazadas, mentres que outras se teme que xa estean extinguidas.[194] As especies de Islandia tenden a ser máis susceptibles, igual que Helictopleurus undatus de Madagascar, que se cre que se extinguiron durante o final do século XX.[195] Os conservacionistas intentaron despertar un gusto ou simpatía polos escaravellos utilizando como buques insignia especies como a vacaloura, Lucanus cervus,[196] e os escaravellos cicindélidos. No Xapón o vagalume Genji, Luciola cruciata, é extremadamente popular, e en Suráfrica o escaravello peloteiro elefante Addo poden servir para ampliar o ecotourismo máis alá dos cinco grandes mamíferos turísticos. O desgusto popular polos escaravellos que causan pragas, tamén pode ser aproveitado para crear interese polos insectos, ou as adaptacións ecolóxicas pouco comúns de especies como o escaravello que captura camaróns, Cicinis bruchi.[197]
Galería de imaxes[editar | editar a fonte]
- Artigo principal: Galería de imaxes de escaravellos de Galicia.




Notas[editar | editar a fonte]
- Notas a rodapé
- ↑ Na fórmula tarsal cóntanse os segmentos tarsais da patas anteriores, medias e posteriores; por exemplo, 5-5-4.
- ↑ O xénero de escaravellos Cerambyx foi nomeado así en referencia a el.
- ↑ A placa foi etiquetada como "Neocerambyx æneas, Cladognathus tarandus, Diurus furcellatus, Ectatorhinus Wallacei, Megacriodes Saundersii, Cyriopalpus Wallacei".
- Referencias
- ↑ 1,0 1,1 Bouchard, P.; Bousquet, Y.; Davies, A.; Alonso-Zarazaga, M.; Lawrence, J.; Lyal, C.; Newton, A.; Reid, C.; Schmitt, M.; Ślipiński, A.; Smith, A. (2011). "Family-group names in Coleoptera (Insecta)". ZooKeys 88 (88): 1–972. PMC 3088472. PMID 21594053. doi:10.3897/zookeys.88.807.
- ↑ Definicións no Dicionario da Real Academia Galega e no Portal das Palabras para coleópteros.
- ↑ Definicións no Dicionario da Real Academia Galega e no Portal das Palabras para escaravello.
- ↑ 4,0 4,1 Definición de escaravello no Dicionario de Galego de Ir Indo e a Xunta de Galicia.
- ↑ Diccionario etimológico coleóptero
- ↑ Etimologías de Chile escarabajo
- ↑ 7,0 7,1 7,2 7,3 7,4 7,5 7,6 7,7 7,8 Gilliott, Cedric (August 1995). Entomology (2 ed.). Springer-Verlag. p. 96. ISBN 0-306-44967-6.
- ↑ 8,00 8,01 8,02 8,03 8,04 8,05 8,06 8,07 8,08 8,09 8,10 8,11 8,12 8,13 8,14 8,15 8,16 McHugh (2009)
- ↑ Rosenzweig, M. L. (1995). Species Diversity in Space and Time. Cambridge: Cambridge University Press. p. 2. ISBN 978-0-521-49952-1.
- ↑ 10,0 10,1 Hunt, T.; Bergsten, J.; Levkanicova, Z.; Papadopoulou, A.; John, O. S.; Wild, R.; Hammond, P. M.; Ahrens, D.; Balke, M.; Caterino, M. S.; Gómez-Zurita, J.; Ribera, I; Barraclough, T. G.; Bocakova, M.; Bocak, L; Vogler, A. P. (2007). "A Comprehensive Phylogeny of Beetles Reveals the Evolutionary Origins of a Superradiation". Science 318 (5858): 1913–1916. Bibcode:2007Sci...318.1913H. PMID 18096805. doi:10.1126/science.1146954.
- ↑ Hammond, P.M. (1992). Species inventory. Global Biodiversity, Status of the Earth's Living Resources: a Report (1st ed.) (London: Chapman & Hall). pp. 17–39. ISBN 978-0-412-47240-4.
- ↑ 12,0 12,1 Stork, Nigel E.; McBroom, James; Gely, Claire; Hamilton, Andrew J. (2015). "New approaches narrow global species estimates for beetles, insects, and terrestrial arthropods". PNAS 116 (24): 7519–7523. Bibcode:2015PNAS..112.7519S. PMC 4475949. PMID 26034274. doi:10.1073/pnas.1502408112.
- ↑ Gullan, P.J.; Cranston, P.S. (2014). The Insects: An Outline of Entomology (5 ed.). John Wiley & Sons. p. 6. ISBN 1-4443-3036-5.
- ↑ Gullan, P. J.; Cranston, P. S. (2014). The Insects: An Outline of Entomology (5 ed.). John Wiley & Sons. p. 517. ISBN 1-4443-3036-5.
- ↑ "Heaviest insect". Guinness World Records. Consultado o 2017-02-01.
- ↑ Williams, David M. (2001). "Chapter 30 — Largest Insect". Book of Insect Records. Universidade de Florida. Arquivado dende o orixinal o 18 de xullo de 2011. Consultado o 02 de xullo de 2018.
- ↑ Polilov, Alexey (2015). "How small is the smallest? New record and remeasuring of Scydosella musawasensis Hall, 1999 (Coleoptera, Ptiliidae), the smallest known free-living insect". ZooKeys 526: 61–64. PMC 4607844. PMID 26487824. doi:10.3897/zookeys.526.6531.
- ↑ 18,0 18,1 18,2 18,3 18,4 18,5 18,6 18,7 Benisch, Christoph (2010). "Phylogeny of the beetles". The beetle fauna of Germany. Kerbtier. Consultado o March 16, 2011.
- ↑ Grimaldi, David; Engel, Michael S. (2005). Evolution of the Insects. Cambridge University Press. p. 397. ISBN 978-1-107-26877-7.
Os Curculionoidea triásicos teñen o mesmo status que os Chrysoleloidea triásicos: unha relación que é altamente improbable e actualemtne imposible de verificar.
- ↑ Béthoux, Oliver (2009). "The earliest beetle identified". Journal of Paleontology 83 (6): 931–937. doi:10.1666/08-158.1.
- ↑ Hörnschemeyer, T.; Stapf, H. "Die Insektentaphozönose von Niedermoschel (Asselian, unt. Perm; Deutschland)". Schriften der Alfred-Wegener-Stiftung (en German) (99/8): 98.
- ↑ Kirejtshuk, Alexander G.; Poschmann, Markus; Prokop, Jakub; Garrouste, Romain; Nel, André (2014-07-04). "Evolution of the elytral venation and structural adaptations in the oldest Palaeozoic beetles (Insecta: Coleoptera: Tshekardocoleidae)". Journal of Systematic Palaeontology (en inglés) 12 (5): 575–600. ISSN 1477-2019. doi:10.1080/14772019.2013.821530.
- ↑ Kukalová, J. (1969). "On the systematic position of the supposed Permian beetles, Tshecardocoleidae, with a description of a new collection from Moravia". Sborník geologických Věd, Paleontologie 11: 139–161.
- ↑ Beckemeyer, R. J.; Engel, M. S. (2008). "A second specimen of Permocoleus (Coleoptera) from the Lower Permian Wellington Formation of Noble County, Oklahoma" (PDF). Journal of the Kansas Entomological Society 81 (1): 4–7. doi:10.2317/JKES-708.01.1. Arquivado dende o orixinal (PDF) o 18 de xullo de 2011. Consultado o 02 de xullo de 2018.
- ↑ Shcherbakov, D. E. (2008). "On Permian and Triassic insect faunas in relation to biogeography and the Permian-Triassic crisis". Paleontological Journal 42 (1): 15–31. doi:10.1007/s11492-008-1003-1.
- ↑ Ponomarenko, A. G. (2004). "Beetles (Insecta, Coleoptera) of the Late Permian and Early Triassic" (PDF). Paleontological Journal 38 (Suppl. 2): S185–S196. Arquivado dende o orixinal (PDF) o 11 de novembro de 2013. Consultado o 02 de xullo de 2018.
- ↑ 27,0 27,1 27,2 McHugh (2009), p. 186
- ↑ Labandeira, C. C.; Sepkoski, J. J. (1993). "Insect diversity in the fossil record" (PDF). Science 261 (5119): 310–315. Bibcode:1993Sci...261..310L. PMID 11536548. doi:10.1126/science.11536548. Arquivado dende o orixinal (PDF) o 31 de marzo de 2012. Consultado o 02 de xullo de 2018.
- ↑ Gratshev, Vadim G.; Zherikhin, Vladimir V. (15 de outubro de 2003). "Insect diversity in the fossil record" (PDF). Acta Zoologica Cracoviensia (Fossil Insects) 261 (5119): 129–138. Bibcode:1993Sci...261..310L. PMID 11536548. doi:10.1126/science.11536548. Arquivado dende o orixinal (PDF) o 29 de decembro de 2016. Consultado o 02 de xullo de 2018.
- ↑ Chang, H.; Zhang, F.; Ren, D. (2008). "A new genus and two new species of fossil elaterids from the Yixian Formation of Western Liaoning, China (Coleoptera: Elateridae)" (PDF). Zootaxa (1785): 54–62. Arquivado dende o orixinal (PDF) o 04 de xullo de 2011. Consultado o 02 de xullo de 2018.
- ↑ Orekhovo-Zuyevo, A. V. A. (1993). "Jurassic and Lower Cretaceous Buprestidae (Coleoptera) from Eurasia" (PDF). Paleontological Journal (1A): 9–34.
- ↑ Ponomarenko, Alexandr G. (1985). "Fossil insects from the Tithonian 'Solnhofener Plattenkalke' in the Museum of Natural History, Vienna" (PDF). Annalen des Naturhistorischen Museums in Wien 87 (1): 135–144.
- ↑ Yan, E. V. (2009). "A new genus of elateriform beetles (Coleoptera, Polyphaga) from the Middle-Late Jurassic of Karatau" (PDF). Paleontological Journal 43 (1): 78–82. doi:10.1134/S0031030109010080. Arquivado dende o orixinal (PDF) o 18 de xullo de 2011. Consultado o 02 de xullo de 2018.
- ↑ 34,0 34,1 Tan, J.-J.; Ren, D.; Liu, M. (2005). "New ommatids from the Late Jurassic of western Liaoning, China (Coleoptera: Archostemata)" (PDF). Insect Science 12 (3): 207–216. doi:10.1111/j.1005-295X.2005.00026.x. Arquivado dende o orixinal (PDF) o 18 de xullo de 2011. Consultado o 02 de xullo de 2018.
- ↑ Ponomarenko, A. G. (1997). "New beetles of the family Cupedidae from the Mesozoic of Mongolia. Ommatini, Mesocupedini, Priacmini" (PDF). Paleontological Journal 31 (4): 389–399. Arquivado dende o orixinal (PDF) o 2006-09-25.
- ↑ Alexeev, A. V. (2009). "New Jewel Beetles (Coleoptera: Buprestidae) from the Cretaceous of Russia, Kazakhstan, and Mongolia" (PDF). Paleontological Journal 43 (3): 277–281. doi:10.1134/s0031030109030058. Arquivado dende o orixinal (PDF) o 18 de xullo de 2011. Consultado o 02 de xullo de 2018.
- ↑ Chin, K.; Gill, B. D. (1996). "Dinosaurs, dung beetles, and conifers; participants in a Cretaceous food web". Palaois 11 (3): 280–285. Bibcode:1996Palai..11..280C. doi:10.2307/3515235.
- ↑ Arillo, Antonio & Ortuño, Vicente M. (2008). "Did dinosaurs have any relation with dung-beetles? (The origin of coprophagy)". Journal of Natural History 42 (19&20): 1405–1408. doi:10.1080/00222930802105130.
- ↑ McNamara, M. E.; Briggs, D. E. G.; Orr, P. J.; Noh, H.; Cao, H. (2011). "The original colours of fossil beetles". Proceedings of the Royal Society B: Biological Sciences 279 (1731): 1114. PMC 3267148. doi:10.1098/rspb.2011.1677.
- ↑ Coope, G. R. (1979). "Late Cenozoic Fossil Coleoptera: Evolution, Biogeography, and Ecology". Annual Review of Ecology and Systematics 10: 246–267. JSTOR 2096792.
- ↑ Hutchinson, G. E. (1959). "Homage to Santa Rosalia or why are there so many kinds of animals?". The American Naturalist 93 (870): 145–159. JSTOR 2458768. doi:10.1086/282070.
- ↑ Maddison, D. R. (1995). "Polyphaga". Tree of Life web project. Arquivado dende o orixinal o 07 de marzo de 2016. Consultado o 27 February 2016.
- ↑ Beutel, R. G.; Lawrence, J. F. (2005). "4. Coleoptera (Morphology)". En Kristensen, N. P.; Beutel, R. G. Handbook of Zoology, Band 4: Arthropoda, 2: Insecta, Coleoptera, Beetles. Volume 1: Morphology and Systematics (Archostemata, Adephaga, Myxophaga, Polyphaga partim). p. 23. ISBN 3-11-017130-9.
- ↑ Beutel, R. G.; Ribera, I. (2005). "7. Adephaga Schellenberg, 1806". En Kristensen, N. P.; Beutel, R. G. Handbook of Zoology, Band 4: Arthropoda, 2: Insecta, Coleoptera, Beetles. Volume 1: Morphology and Systematics (Archostemata, Adephaga, Myxophaga, Polyphaga partim). p. 54. ISBN 3-11-017130-9.
- ↑ "Suborder Archostemata – Reticulated and Telephone-pole Beetles". BugGuide. 2006. Consultado o 2017-01-26.
- ↑ 46,0 46,1 46,2 Lawrence, John F.; Ślipiński, Adam (2013). Australian Beetles. Morphology, Classification and Keys. CSIRO. pp. 1–16. ISBN 978-0-643-09728-5.
- ↑ Mesaros, Gabor (2013). "Sphaeriusidae (Coleoptera, Myxophaga): A new beetle family to the fauna of Serbia". Bulletin of the Natural History Museum (6): 71–74. doi:10.5937/bnhmb1306071m.
- ↑ Beutel, Rolf G.; Leschen, Richard (2005). Handbook of Zoology. Volume 4. Part 38. Arthropoda. Morphology and Systematics (Archostemata, Adephaga, Myxophaga, Polyphaga partim). Walter de Gruyter. p. 43.
- ↑ Whiting, Michael F. (2002). "Phylogeny of the holometabolous insect orders: molecular evidence". Zoologica Scripta 31 (1): 3–15. doi:10.1046/j.0300-3256.2001.00093.x.
- ↑ Beutel, R.; Haas, F. (2000). "Phylogenetic relationships of the suborders of Coleoptera (Insecta)". Cladistics 16: 103–141. doi:10.1111/j.1096-0031.2000.tb00350.x.
- ↑ 51,0 51,1 Kukalová-Peck, J.; Lawrence, J. F. (1993). "Evolution of the hind wing in Coleoptera". Canadian Entomologist 125 (2): 181–258. doi:10.4039/Ent125181-2.
- ↑ Maddison, D. R.; Moore, W.; Baker, M. D.; Ellis, T. M.; Ober, K. A.; Cannone, J. J.; Gutell, R. R. (2009). "Monophyly of terrestrial adephagan beetles as indicated by three nuclear genes (Coleoptera: Carabidae and Trachypachidae)". Zoologica Scripta 38 (1): 43–62. PMC 2752903. PMID 19789725. doi:10.1111/j.1463-6409.2008.00359.x.
- ↑ Maddison, David R. (September 11, 2000). "Coleoptera. Beetle". Tree of Life Web Project. tolweb.org. Arquivado dende o orixinal o 17 de maio de 2022. Consultado o March 18, 2011.
- ↑ Niehuis, Oliver; Hartig, Gerrit; Grath, Sonja; et al. (2012). "Genomic and Morphological Evidence Converge to Resolve the Enigma of Strepsiptera". Current Biology 22 (14): 1309–1313. PMID 22704986. doi:10.1016/j.cub.2012.05.018.
- ↑ 55,0 55,1 55,2 McKenna, Duane D.; Wild, Alexander L.; et al. (2015). "The beetle tree of life reveals that Coleoptera survived end-Permian mass extinction to diversify during the Cretaceous terrestrial revolution". Systematic Entomology 40 (4): 835–880. doi:10.1111/syen.12132.
- ↑ Hunt, Toby; et al. (2007). "A Comprehensive Phylogeny of Beetles Reveals the Evolutionary Origins of a Superradiation". Science 318 (5858): 1913–19116. Bibcode:2007Sci...318.1913H. PMID 18096805. doi:10.1126/science.1146954.
- ↑ "Introduction to the Identification of Beetles (Coleoptera)" (PDF). Universidade de Florida. Consultado o 15 March 2017.
- ↑ 58,0 58,1 Hangay, G.; Zborowski, P. (2010). A Guide to the Beetles of Australia. CSIRO. p. 10.
- ↑ 59,0 59,1 Christoph Benisch (2007). "3. Antennae, Beetle morphology". Kerbtier.de (Beetle fauna of Germany). Consultado o 2017-03-14.
- ↑ Zurek, D. B.; Gilbert, C. (2014). "Static antennae act as locomotory guides that compensate for visual motion blur in a diurnal, keen-eyed predator". Proceedings of the Royal Society B: Biological Sciences 281 (1779): 20133072. PMC 3924084. doi:10.1098/rspb.2013.3072.
- ↑ Berkov, Amy; Rodríguez, Nelson; Centeno, Pedro (2007). "Convergent evolution in the antennae of a cerambycid beetle, Onychocerus albitarsis, and the sting of a scorpion". Naturwissenschaften 95 (3): 257. Bibcode:2008NW.....95..257B. PMID 18004534. doi:10.1007/s00114-007-0316-1.
- ↑ "Antennae Types". University of Sydney. Arquivado dende o orixinal o 22 de febreiro de 2018. Consultado o 2017-01-26.
- ↑ Arnett, Jr., R. H.; Thomas, M. C. (2001). American Beetles, Volume I: Archostemata, Myxophaga, Adephaga, Polyphaga: Staphyliniformia. CRC Press. pp. 3–7. ISBN 1-4822-7432-9.
- ↑ "Anatomy: Head, Thorax, Abdomen, and Genitalia (Insects)". what-when-how.com. Arquivado dende o orixinal o 22 de xuño de 2018. Consultado o 2017-01-26.
- ↑ Green, Kristina Karlsson; Kovalev, Alexander; Svensson, Erik I.; Gorb, Stanislav N. (2013). "Male clasping ability, female polymorphism and sexual conflict: fine-scale elytral morphology as a sexually antagonistic adaptation in female diving beetles". Journal of The Royal Society Interface 10 (86): 20130409. PMC 3730688. PMID 23825114. doi:10.1098/rsif.2013.0409.
- ↑ Burkness, S.; Hahn, J. (2007). "Flea beetles in home gardens". University of Minnesota (Extension). Consultado o 2017-01-26.
- ↑ 67,0 67,1 Carpenter, George Herbert (1899). Insects, their structure and life.
- ↑ Philips, Chris; Fread, Elizabeth; Kuhar, Tom. "Leatherwing (Soldier) Beetles" (PDF). Arquivado dende o orixinal (PDF) o 30 de novembro de 2016. Consultado o 14 March 2017.
- ↑ Donald W. Hall; Marc A. Branham (2016). "Calopteron discrepans (Newman) (Insecta: Coleoptera: Lycidae)". Universidade de Florida. Consultado o 2017-03-15.
- ↑ Lawrence, J. F.; Hastings, A. M.; Dallwitz, M. J.; Paine, T. A.; Zurcher, E. J. (2005). "Elateriformia (Coleoptera): descriptions, illustrations, identification, and information retrieval for families and subfamilies". Arquivado dende o orixinal o 21 de outubro de 2018. Consultado o 2017-01-26.
- ↑ Beenen, R; Winkelman, J. K. (2001). "Aantekeningen over Chrysomelidae in Nederland 5". Entomologische Berichten (en Dutch) 61: 63–67.
- ↑ Arnett, Ross H., Jr; Thomas, Michael C. (2000). American Beetles, Volume I: Archostemata, Myxophaga, Adephaga, Polyphaga: Staphyliniformia. CRC Press. p. 8. ISBN 978-1-4822-7432-5.
- ↑ Seymour, Roger S.; Matthews, Philip G. D. (2012). "Physical gills in diving insects and spiders: Theory and experiment". The Journal of Experimental Biology 216 (2): 164. doi:10.1242/jeb.070276.
- ↑ Schmidt-Nielsen, Knut (January 15, 1997). "Insect Respiration". Animal Physiology: Adaptation and Environment (5th ed.). Cambridge University Press. p. 55. ISBN 0-521-57098-0.
- ↑ Miller, T. A. (1985). "Chapter 8: Structure and Physiology of the Circulatory System". En Kerkut, G. A.; Gilbert, L. I. Comprehensive Insect Physiology, Biochemistry and Pharmacology. Volume 3: Integument, Respiration and Circulation. Pergamom Press. pp. 289–355. ISBN 0-08-030804-X.
- ↑ Scoble, M. J. (1992). The Lepidoptera: Form, function, and diversity. Oxford University Press. ISBN 978-1-4020-6242-1.
- ↑ Ozbek, H.; Szalokia, D. (1998). "A contribution to the knowledge of the Meloidae (Coleoptera) fauna of Turkey along with new record". Turkish Journal of Zoology 22: 23–40.
- ↑ Medvedev, L. N.; Pavlov, S. I. (1988). "Mating behavior of the Chrysomelidae (Coleoptera)". Entomological Review 67: 100–108.
- ↑ Crowson, R. A. (2013). The Biology of the Coleoptera. Elsevier Science. pp. 358–370. ISBN 978-1-4832-1760-4.
- ↑ Beck, S. D.; Bharadwaj, R. K. (1972). "Reversed development and cellular ageing in an insect". Science 178 (4066): 1210–1211. Bibcode:1972Sci...178.1210B. PMID 4637808. doi:10.1126/science.178.4066.1210.
- ↑ "Definition of 'Scarabaeiform'". Amateur Entomologists' Society. Consultado o 2017-01-27.
- ↑ Krinsky, W. L. (2009). "8 Beetles (Coleoptera)". En Mullen, G. R.; Durden, L. A. Medical and Veterinary Entomology (2nd ed.). Elsevier. pp. 101–115. ISBN 978-0-12-372500-4.
- ↑ "Hypermetamorphosis of Striped Blister Beetle – Epicauta vittata". BugGuide. 2007. Consultado o 2017-01-27.
- ↑ Zeng, Yong (1995). "Longest Life Cycle". Universidade de Florida. Consultado o 17 March 2017.
- ↑ Jeffries, Daniel L.; Chapman, Jason; Roy, Helen E.; Humphries, Stuart; Harrington, Richard; Brown, Peter M. J.; Handley, Lori-J. Lawson (2013). "Characteristics and Drivers of High-Altitude Ladybird Flight: Insights from Vertical-Looking Entomological Radar". PLOS ONE 8 (2): e82278. Bibcode:2013PLoSO...882278J. doi:10.1371/journal.pone.0082278.
- ↑ Capinera, John L. (2008). Encyclopedia of Entomology (2 ed.). Springer Science & Business Media. pp. 3218–3219. ISBN 978-1-4020-6242-1.
- ↑ Arnett R. H., Jr.; Thomas, M. C. (2001). "Haliplidae". American Beetles, Volume 1. CRC Press. pp. 138–143. ISBN 0-8493-1925-0.
- ↑ "Mountain Pine Beetle – Beetle Love". Parks Canada. Consultado o March 13, 2011.
- ↑ Wyatt, T. D. & Foster, W. A. (1989). "Parental care in the subsocial intertidal beetle, Bledius spectabilis, in relation to parasitism by the ichneumonid wasp, Barycnemis blediator". Behaviour 110 (1–4): 76–92. JSTOR 4534785. doi:10.1163/156853989X00394.
- ↑ Milne, Lorus J.; Milne, Margery J. (1944). "Notes on the Behavior of Burying Beetles (Nicrophorus spp.)". Journal of the New York Entomological Society 52 (4): 311–327. JSTOR 25005075.
- ↑ Hanski, Ilkka; Yves, Cambefort (1991). Dung Beetle Ecology. Princeton University Press. pp. 626–672. ISBN 0-691-08739-3.
- ↑ "Beetle | San Diego Zoo Animals & Plants". animals.sandiegozoo.org (en inglés). Consultado o 2017-07-18.
- ↑ Crespi, B. J.; Yanega, D. (1995). "The definition of eusociality". Behavioral Ecology 6 (1): 109–115. doi:10.1093/beheco/6.1.109.
- ↑ 94,0 94,1 Kent, D. S. & Simpson, J. A. (1992). "Eusociality in the beetle Austroplatypus incompertus (Coleoptera: Curculionidae)". Naturwissenschaften 79 (2): 86–87. Bibcode:1992NW.....79...86K. doi:10.1007/BF01131810.
- ↑ 95,0 95,1 "Science: The Australian beetle that behaves like a bee". New Scientist. 1992-05-09. Consultado o 2010-10-31.
- ↑ Schuster, Jack C.; Schuster, Laura B. (1985). "Social behavior in Passalid beetles (Coleoptera: Passalidae): Cooperative brood care". Florida Entomologist 68 (2): 266–272. doi:10.2307/3494359. Arquivado dende o orixinal o 02 de marzo de 2016. Consultado o 02 de xullo de 2018.
- ↑ Lobanov, A.L. (2002). "feeding". Beetle Biology And Ecology. Beetles (Coleoptera) and Coleopterologist. Consultado o March 13, 2011.
- ↑ Islam, M.; Hossain, A.; Mostafa, M. G.; Hossain, M. M. (2016). "Forensically important insects associated with the decomposition of mice carrion in Bangladesh". Jahangirnagar University Journal of Biological Science 5 (1): 11–20. doi:10.3329/jujbs.v5i1.29739.
- ↑ Grebennikov, Vasily V.; Leschen, Richard A. B. (2010). "External exoskeletal cavities in Coleoptera and their possible mycangial functions". Entomological Science 13 (1): 81–98. doi:10.1111/j.1479-8298.2009.00351.x.
- ↑ 100,0 100,1 100,2 Evans & Bellamy (2000), pp. 27–28
- ↑ Cott, H. B. (1940). Adaptive Coloration in Animals. Methuen. p. 414.
- ↑ 102,0 102,1 102,2 Evans & Bellamy (2000), p. 126
- ↑ 103,0 103,1 Evans & Bellamy (2000)
- ↑ McHugh (2009), p. 199
- ↑ Peck, Stewart B. (2006). "Distribution and biology of the ectoparasitic beaver beetle Platypsyllus castoris Ritsema in North America (Coleoptera: Leiodidae: Platypsyllinae)". Insecta Mundi 20 (1–2): 85–94.
- ↑ Neumann, P. & Elzen, P. J. (2004). "The biology of the small hive beetle (Aethina tumida, Coleoptera: Nitidulidae): Gaps in our knowledge of an invasive species". Apidologie 35 (3): 229–247. doi:10.1051/apido:2004010. Arquivado dende o orixinal o 22 de setembro de 2017. Consultado o 02 de xullo de 2018.
- ↑ Meyer, John R. (March 8, 2005). "Coleoptera". North Carolina State University. Arquivado dende o orixinal o 06 de marzo de 2016. Consultado o March 13, 2011.
- ↑ Weber, Donald C.; Saska, Pavel; Chaboo, Caroline S. (2008). "Carabid Beetles (Coleoptera: Carabidae) as Parasitoids". Encyclopedia of Entomology. Springer Netherlands. pp. 719–721. doi:10.1007/978-1-4020-6359-6_492.
- ↑ Gullan, P. J.; Cranston, P. S. (2014). The Insects: An Outline of Entomology (5 ed.). Wiley, John & Sons. p. 314. ISBN 1-4443-3036-5.
- ↑ Jones, G. D. & Jones, S. D. (2001). "The uses of pollen and its implication for Entomology". Neotropical Entomology 30 (3): 314–349. doi:10.1590/S1519-566X2001000300001.
- ↑ Ollerton, J.; Johnson, S. D.; Cranmer, L. & Kellie, S. (2003). "The pollination ecology of an assemblage of grassland asclepiads in South Africa". Annals of Botany 92 (6): 807–834. PMC 4243623. PMID 14612378. doi:10.1093/aob/mcg206.
- ↑ 112,0 112,1 Malloch, D.; Blackwell, M. (1993). "Dispersal biology of ophiostomatoid fungi". En Wingfield, M. J.; K. A. Seifert; J. F. Webber. Ceratocystis and Ophiostoma: Taxonomy, Ecology and Pathology. St. Paul: APS. pp. 195–206. ISBN 978-0-89054-156-2.
- ↑ Scott, J. J.; Oh, D. C.; Yuceer, M. C.; Klepzig, K. D.; Clardy, J.; Currie, C. R. (2008). "Bacterial protection of beetle-fungus mutualism". Science 322 (5898): 63. Bibcode:2008Sci...322...63S. PMC 2761720. PMID 18832638. doi:10.1126/science.1160423.
- ↑ Francke-Grossmann, H. (1967). "Ectosymbiosis in wood inhabiting insects". En M. Henry. Symbiosis 2. New York: Academic Press. pp. 141–205.
- ↑ Ivo Hodek (2012). "Review Article: Adult Diapause in Coleoptera". Psyche: A Journal of Entomology 2012: 1. doi:10.1155/2012/249081.
- ↑ Kaufmann, T. (1971). "Hibernation in the Arctic Beetle, Pterostichus brevicornis, in Alaska". Journal of the Kansas Entomological Society 44 (1): 81–92.
- ↑ Gullan, P. J.; Cranston, P. S. (1994). The Insects: An Outline of Entomology. Chapman and Hall. pp. 103–104. ISBN 0-412-49360-8.
- ↑ Lombadero, Maria J.; Ayres, Matthew P.; Ayres, Bruce D.; Reeve, John D. (2000). "Cold Tolerance of Four Species of Bark Beetle (Coleoptera: Scolytidae) in North America" (PDF). Environmental Ecology 29 (3): 421–432.
- ↑ Sformo, T.; Walters, K.; Jeannet, K.; Wowk, B.; Fahy, G. M.; Barnes, B. M.; Duman, J. G. (2010). "Deep supercooling, vitrification and limited survival to −100 °C in the Alaskan beetle Cucujus clavipes puniceus (Coleoptera: Cucujidae) larvae". Journal of Experimental Biology 213 (3): 502–509. PMID 20086136. doi:10.1242/jeb.035758.
- ↑ Christopher Brooks (2013-03-26). "The life of extremophiles: Surviving in hostile habitats". BBC Nature. Arquivado dende o orixinal o 08 de xullo de 2018. Consultado o 2017-03-16.
- ↑ Graham, L. A; Liou, Y. C.; Walker, V. K.; Davies, P. L. (August 1997). "Hyperactive antifreeze protein from beetles". Nature 388 (6644): 727–728. PMID 9285581. doi:10.1038/41908.
O escaravello da fariña, Tenebrio molitor, contén unha familia de pequenas proteínas de histérese térmica ricas en Cys e Thr que rebaixan o punto de conxelación da hemolinfa ata en 5,5 °C por debaixo do punto de conxelación (ΔT=histérese térmica). A expresión dunha proteína de histérese térmica foi avaliada ao longo do desenvolvemento e despois da exposición a condicións ambientais alteradas.
- ↑ Walters, K. R. Jr; Serianni, A. S.; Sformo, T.; Barnes, B. M.; Duman, J. G. (2009). "A nonprotein thermal hysteresis-producing xylomannan antifreeze in the freeze-tolerant Alaskan beetle Upis ceramboides". PNAS 106 (48): 20210–20215. Bibcode:2009PNAS..10620210W. PMC 2787118. PMID 19934038. doi:10.1073/pnas.0909872106.
- ↑ Walters, Jr., K. R.; Pan, Q.; Serianni, A. S.; Duman, J. G. (2009). "Cryoprotectant biosynthesis and the selective accumulation of threitol in the freeze-tolerant Alaskan beetle, Upis ceramboides". Journal of Biological Chemistry 284 (25): 16822–16831. PMC 2719318. PMID 19403530. doi:10.1074/jbc.M109.013870.
- ↑ Edney, E. B. (1971). "The Body Temperature of Tenerionid Beetles in the Namid Desert of Southern Africa" (PDF). Journal of Experimental Biology 55: 253–272.
- ↑ Knisley, C. B.; Schultz, T. D.; Hasewinkel, T. H. (1990). "Seasonal Activity and Thermoregulatory Behavior of Cicindela patruela (Coleoptera: Cicindelidae)". Annals of the Entomological Society of America 83 (5): 911. doi:10.1093/aesa/83.5.911.
- ↑ Parker, Andrew R.; Lawrence, Chris R. (2001-11-01). "Water capture by a desert beetle". Nature 414 (6859): 33–34. Bibcode:2001Natur.414...33P. PMID 11689930. doi:10.1038/35102108. Consultado o 2017-03-14.
- ↑ Hoback, W. Wyatt; Stanley, David W.; Higley, Leon G.; Barnhart, M. Christopher. "Survival of Immersion and Anoxia by Larval Tiger Beetles, Cicindela togata". The American Midland Naturalist 140 (1): 27–33. doi:10.1674/0003-0031(1998)140[0027:SOIAAB]2.0.CO;2.
- ↑ Conradi-Larsen, Else-Margrete; Sømme, Lauritz (1973). "Anaerobiosis in the Overwintering Beetle Pelophila borealis". Nature 245: 388–390. Bibcode:1973Natur.245..388C. doi:10.1038/245388a0.
- ↑ Allison, R. (2015-03-17). "Half of the UK sees start of pollen beetle migration". Consultado o 2017-03-16.
- ↑ Southwood, T. R. E. (1962). "Migration of Terrestrial Arthropods in Relation to Habitat". Biological Reviews 37 (2): 171. doi:10.1111/j.1469-185X.1962.tb01609.x.
- ↑ Dingle, H. (2014). Migration: The Biology of Life on the Move. Oxford University Press.
- ↑ 132,0 132,1 Zabludoff, Marc (2008). Beetles. Malaysia: Michelle Bison. pp. 14–17. ISBN 978-0-7614-2532-8.
- ↑ Cambefort, Yves (2011). "Beetles as religious symbols". Insects.org. Arquivado dende o orixinal o 18 de outubro de 2016. Consultado o 10 February 2017.
- ↑ Dollinger, André (January 2002). "Ancient Egyptian bestiary: Insects". Arquivado dende o orixinal o 01 de abril de 2015. Consultado o July 19, 2011.
- ↑ Morales-Correa, Ben (2006). "Egyptian Symbols". All-About-Egypt. Arquivado dende o orixinal o 20 de agosto de 2011. Consultado o July 19, 2011.
- ↑ Ussishkin, David (2004). The New Archaeological Excavations at Lachish (1973–1994). Tel Aviv: Institute of Archaeology of Tel Aviv University. Arquivado dende o orixinal o 03 de xullo de 2018. Consultado o 02 de xullo de 2018.
- ↑ "Isis and Osiris", Moralia, no volume V da Loeb Classical Library, 1936. "A raza dos escaravellos non ten femias, pero todos os machos exectan esperma nun paquete redondo de material que fan rodar empuxándoo desde o lado oposto, xusto igual a como oo sol parece xirar nos ceos na dirección oposta ao seu propio curso, que é de oeste a leste.
- ↑ Betz, H. D., ed. (1992). The Greek Magical Papyri in Translation (Including the Demotic Spells) (2nd ed.). University of Chicago Press. pp. IV.52–85; VII.520; XII.101; XIII.1065–1070; XXXVI.170.
- ↑ Pliny the Elder. "Natural History Book 11". Perseus.tufts.edu. Consultado o 2017-01-25.
- ↑ Pliny the Elder. "Natural History Book 11, Chapter 34". Perseus.tufts.edu. Consultado o 2017-01-25.
- ↑ 141,0 141,1 Beavis, I. C. (1988). Insects and other Invertebrates in Classical Antiquity. University of Exeter. pp. 153–154.
- ↑ Antonino Liberalis. Metamorphoses. Trans. Celoria, F. 1992. The Metamorphoses of Antoninus Liberalis: A translation with a commentary. London and New York, Routledge.
- ↑ Sprecher-Uebersax, E. (2008). "The Stag Beetle Lucanus Cervus (Coleoptera, Lucanidae) in Art and Mythology". Revue d' Ecologie 63: 145–151.
- ↑ Eladio Rodríguez González (1958-1961): Diccionario enciclopédico gallego-castellano, Galaxia, Vigo http://sli.uvigo.es/ddd/ddd_pescuda.php?pescuda=escarabello&tipo_busca=lema
- ↑ 145,0 145,1 Mississippi State University. "History of the Boll Weevil in the United States". Economic impacts of the boll weevil. Arquivado dende o orixinal o May 12, 2008.
- ↑ 146,0 146,1 University of Florida (2007). "Featured creatures: Leptinotarsa spp". Retrieved 2017-03-19.
- ↑ "Species Leptinotarsa decemlineata - Colorado Potato Beetle". BugGuide. 2017. Consultado o 2017-03-19..
- ↑ "Invasion history: Leptinotarsa decemlineata, Colorado Beetle". Non-Native Species Secretariat (DEFRA). 2017. Arquivado dende o orixinal o 28 de xullo de 2020. Consultado o 2017-03-20.
- ↑ Lehmann, P. "The Colorado potato beetle is the grandmaster of adaptation". University of Jyväskylä. Arquivado dende o orixinal o 10 de maio de 2017. Consultado o 12 de outubro de 2017.
- ↑ "The Colorado beetle". Finnish Food Safety Authority. 2016. Arquivado dende o orixinal o 08 de xullo de 2018. Consultado o 12 de outubro de 2017.
- ↑ Alyokhin, A.; Baker, M.; Mota-Sanchez, D.; Dively, G.; Grafius, E. (2008). "Colorado potato beetle resistance to insecticides". American Journal of Potato Research 85 (6): 395–413. doi:10.1007/s12230-008-9052-0.
- ↑ Lockwood, Jeffrey A. (2007-10-21). "Bug bomb:Why our next terrorist attack could come on six legs". Boston Globe. Consultado o 2017-02-13.
- ↑ Heather, Neil W.; Hallman, Guy J. (2008). Pest Management and Phytosanitary Trade Barriers. CABI. pp. 17–18. ISBN 1-84593-343-5. doi:10.1079/9781845933432.0000.
- ↑ Allen, E. A.; Humble, L. M. (2002). "Nonindigenous species introductions: A threat to Canada's forests and forest economy". Canadian Journal of Plant Pathology 24 (2): 103. doi:10.1080/07060660309506983.
- ↑ Webber, Joan F. (2000). "Insect Vector Behavior and the Evolution of Dutch Elm Disease". The Elms. p. 47. ISBN 978-1-4613-7032-1. doi:10.1007/978-1-4615-4507-1_3.
- ↑ Adcock, Edward (2005). "Pests – Death watch beetle". Conservation and collective care. University of Oxford. Arquivado dende o orixinal o 10 de xullo de 2011. Consultado o July 17, 2011.
- ↑ Takano, Shun-Ichiro; Takasu, Keiji; Fushimi, Tsutomu; Ichiki, Ryoko T.; Nakamura, Satoshi (2014). "Suitability of four palm species for the development of the invasive pest Brontispa longissima(Coleoptera: Chrysomelidae) in the field". Entomological Science 17 (2): 265. doi:10.1111/ens.12048.
- ↑ "The Mountain Pine Beetle in British Columbia". Natural Resources Canada. August 19, 2008. Arquivado dende o orixinal o 19 de abril de 2010. Consultado o 24 de xuño de 2010.
- ↑ "Insecta: Coleoptera: Coccinellidae". Institute of Food and Agricultural Services, Universidade de Florida. 2014. Consultado o 2017-02-12.
- ↑ "'Deadly ladybird' sighted in UK". BBC News. 5 de outubro de 2004. Consultado o 17 de xuño de 2010.
- ↑ Kromp, B. (1999). "Carabid beetles in sustainable agriculture: a review on pest control efficacy, cultivation aspects and enhancement". Agriculture, Ecosystems and Environment 74 (1–3): 187–228. doi:10.1016/S0167-8809(99)00037-7.
- ↑ "Beetles play an important role in reducing weeds". Rothamsted Research. Arquivado dende o orixinal o 03 de xullo de 2018. Consultado o 14 March 2017.
- ↑ "Zygogramma bicolorata (Mexican beetle)". Invasive Species Compendium. 2012. Consultado o 2017-02-12.
- ↑ Kovalev, O. V.; Reznik, S. Y.; Cherkashin, V. N. (1983). "Specific features of the methods of using Zygogramma Chevr. (Coleoptera, Chrysomelidae) in biological control of ragweeds (Ambrosia artemisiifolia L., A. psilostachya D.C.)". Entomologicheskoe Obozrenije (en Russian) 62: 402–408.
- ↑ 165,0 165,1 Bornemissza, George (1970). "Insectary Studies on the Control of Dung Breeding Flies by the Activity of the Dung Beetle, Onthophagus Gazella F. (Coleoptera: Scarabaeinae)". Australian Journal of Entomology 9: 31–41. doi:10.1111/j.1440-6055.1970.tb00767.x.
- ↑ Brown, Jacqueline; Scholtz, Clarke H.; Janeau, Jean-Louis; Grellier, Seraphine & Podwojewski, Pascal (2010). "Dung beetles (Coleoptera: Scarabaeidae) can improve soil hydrological properties". Applied Soil Ecology 46 (1): 9–16. doi:10.1016/j.apsoil.2010.05.010. hdl:2263/14419.
- ↑ Losey, John E.; Vaughan, Mace (2006). "The economic value of ecological services provided by insects" (PDF). BioScience 56 (4): 311–323. doi:10.1641/0006-3568(2006)56[311:TEVOES]2.0.CO;2. Arquivado dende o orixinal (PDF) o 19 de xuño de 2018. Consultado o 02 de xullo de 2018.
- ↑ Tomberlin, Jeffery K.; Sanford, Michelle R. (2012). "Forensic entomology and wildlife". En Huffman, Jane E.; Wallace, John R. Wildlife Forensics: Methods and Applications. Developments in Forensic Science 6 (2nd ed.). John Wiley & Sons. pp. 81–107. ISBN 978-1-119-95429-3.
- ↑ Dermestid Beetles working on a female Black Bear skull. 2014. Consultado o 2017-01-26.
- ↑ Fernández-Jalvo, Yolanda; Monfort, Maria Dolores Marín (2008). "Experimental taphonomy in museums: Preparation protocols for skeletons and fossil vertebrates under the scanning electron microscopy". Geobios 41: 157. doi:10.1016/j.geobios.2006.06.006.
- ↑ Ramos-Elorduy, Julieta; Menzel, Peter (1998). Creepy crawly cuisine: the gourmet guide to edible insects. Inner Traditions / Bear & Company. p. 5. ISBN 978-0-89281-747-4.
- ↑ Holland, Jennifer S. (14 May 2013). "U.N. Urges Eating Insects; 8 Popular Bugs to Try". National Geographic. Consultado o 26 January 2017.
- ↑ Pearson, David L.; Cassola, Fabio (1992). "World-Wide Species Richness Patterns of Tiger Beetles (Coleoptera: Cicindelidae): Indicator Taxon for Biodiversity and Conservation Studies". Conservation Biology 6 (3): 376–391. JSTOR 2386038. doi:10.1046/j.1523-1739.1992.06030376.x.
- ↑ McGeoch, Melodie A.; Van Rensburg, Berndt J.; Botes, Antoinette (2002). "The verification and application of bioindicators: A case study of dung beetles in a savanna ecosystem". Journal of Applied Ecology 39 (4): 661. doi:10.1046/j.1365-2664.2002.00743.x.
- ↑ Lachat, Thibault; Wermelinger, Beat; Gossner, Martin M.; Bussler, Heinz; Isacsson, Gunnar; Müller, Jörg (2012). "Saproxylic beetles as indicator species for dead-wood amount and temperature in European beech forests". Ecological Indicators 23: 323. doi:10.1016/j.ecolind.2012.04.013.
- ↑ Bohac, Jaroslav (1999). "Staphylinid beetles as bioindicators" (PDF). Agriculture, Ecosystems & Environment 74 (1–3): 357–372. doi:10.1016/S0167-8809(99)00043-2.
- ↑ Life cycle of the rounded jewel beetles, Sternocera spp. วงจรชีวิตของแมลงทับกลมใช้เวลานานถึง 2 ปี – Siam Insect Zoo-Museum. Malaeng.com (2008-10-20). Retrieved on 2013-04-04.
- ↑ Ivie, Michael A. (2002). "105. Zopheridae". En Ross H. Arnett; Michael Charles Thomas. American Beetles: Polyphaga: Scarabaeoidea through Curculionoidea. Volume 2 of American Beetles. CRC Press. pp. 457–462. ISBN 978-0-8493-0954-0.
- ↑ Rennesson, Stephane; Cesard, Nicolas; Grimaud, Emmanuel (2008). "Duels en miniature: la délicate mise en scène des combats de scarabées au nord de la Thaïlande" (PDF). Insectes (en francés) (INRA) 3 (151).
- ↑ 180,0 180,1 Eraldo Medeiros Costa-Neto (2003). "Entertainment with insects: singing and fighting insects around the world. A brief review" (PDF). Etnobiología 3: 21–29. Arquivado dende o orixinal (PDF) o 13 de febreiro de 2017. Consultado o 02 de xullo de 2018.
- ↑ Pemberton, R. W. (1990). "The Korean water beetle game". Pan-Pacific Entomologist 66 (2): 172–174.
- ↑ Bateman, C.; Hulcr, J. (2016). "Predaceous Diving Beetles as Pets and the Self-Cleaning Aquarium" (PDF). Universidade de Florida (IFAS Extension). Consultado o 2017-01-27.
- ↑ Kawahara, A. Y. (2007). "Thirty-foot telescopic nets, bug-collecting video games, and beetle pets: Entomology in modern Japan" (PDF). American Entomologist 53 (3): 160–172. doi:10.1093/ae/53.3.160. Arquivado dende o orixinal (PDF) o 12 de abril de 2016. Consultado o 02 de xullo de 2018.
- ↑ May, Mitchell (11 July 1999). "Yen For Bugs". Chicago Tribune. Consultado o 2017-01-27.
- ↑ Watts, Jonathan (11 August 1999). "Vending machine beetles". The Guardian. Consultado o 2017-01-27.
- ↑ Morse, Deborah Denenholz; Danahay, Martin A. (2007). Victorian Animal Dreams: Representations of Animals in Victorian Literature and Culture. Ashgate Publishing. p. 5. ISBN 978-0-7546-5511-4.
a manía vitoriana de coleccionar escaravellos
- ↑ Wallace, Alfred Russel (1869). The Malay Archipelago: The land of the orang–utan, and the bird of paradise. A narrative of travel, with sketches of man and nature (1 ed.). Macmillan. pp. vii–xiv.
- ↑ Swedish Biomimetics: The μMist Platform Technology (orixinal URL = http://www.swedishbiomimetics.com/biomimetics_folder.pdf) (data de arquivo = 13 de decembro de 2013)
- ↑ "Namib Desert beetle inspires self-filling water bottle". BBC News. 23 November 2012.
- ↑ Emily Singer (2009-01-29). "The Army's Remote-Controlled Beetle". MIT Technology Review. Consultado o 2017-03-16.
- ↑ Feng Cao; Chao Zhang; Hao Yu Choo; Hirotaka Sato (2016). "Insect–computer hybrid legged robot with user-adjustable speed, step length and walking gait". Journal of the Royal Society Interface 13 (116): 1–10. PMC 4843679. doi:10.1098/rsif.2016.0060.
- ↑ Sato, Hirotaka; Doan, Tat Thang Vo; Kolev, Svetoslav; Huynh, Ngoc Anh; Zhang, Chao; Massey, Travis L.; Kleef, Joshua van; Ikeda, Kazuo; Abbeel, Pieter (2015-03-16). "Deciphering the Role of a Coleopteran Steering Muscle via Free Flight Stimulation". Current Biology 25 (6): 798–803. doi:10.1016/j.cub.2015.01.051.
- ↑ Vo Doan, Tat Thang; Tan, Melvin Y.W.; Bui, Xuan Hien; Sato, Hirotaka (2017-11-03). "An Ultralightweight and Living Legged Robot". Soft Robotics. doi:10.1089/soro.2017.0038.
- ↑ Kotze, D. Johan; O'Hara, Robert B. (2003). "Species decline—but why? Explanations of carabid beetle (Coleoptera, Carabidae) declines in Europe". Oecologia 135 (1): 138–148. Bibcode:2003Oecol.135..138K. doi:10.1007/s00442-002-1174-3.
- ↑ Hanski, Ilkka; Koivulehto, Helena; Cameron, Alison; Rahagalala, Pierre (2007). "Deforestation and apparent extinctions of endemic forest beetles in Madagascar". Biology Letters 3 (3): 344–347. PMC 1995085. PMID 17341451. doi:10.1098/rsbl.2007.0043.
- ↑ Campanaro, A.; Zapponi, L.; Hardersen, S.; Méndez, M.; Al Fulaij, N.; Audisio, P.; Bardiani, M.; Carpaneto, G. M.; Corezzola, S.; Della Rocca, F.; Harvey, D.; Hawes, C.; Kadej, M.; Karg, J.; Rink, M.; Smolis, A.; Sprecher, E.; Thomaes, A.; Toni, I.; Vrezec, A.; Zauli, A.; Zilioli, M.; Chiari, S. (2016). "A European monitoring protocol for the stag beetle, a saproxylic flagship species". Insect Conserv Divers 9 (6): 574–584. doi:10.1111/icad.12194.
- ↑ New, T. R. (2009). Beetles in Conservation. John Wiley & Sons. pp. ix, 1–26, passim. ISBN 978-1-4443-1863-0.
Véxase tamén[editar | editar a fonte]
| Wikispecies posúe unha páxina sobre: Coleópteros |
Bibliografía[editar | editar a fonte]
- Evans, Arthur V.; Bellamy, Charles (2000). An Inordinate Fondness for Beetles. University of California Press. ISBN 978-0-520-22323-3.
- McHugh, Joseph V.; Liebherr, James K. (2009). "Coleoptera". En Vincent H. Resh; Ring T. Cardé. Encyclopedia of Insects (2nd ed.). Academic Press. pp. 183–201. ISBN 978-0-12-374144-8.
- Beckmann, Poul. Living Jewels: The Natural Design of Beetles. ISBN 3-7913-2528-0.
- Cooter, J.; M. V. L. Barclay, eds. (2006). A Coleopterist's Handbook. Amateur Entomological Society. ISBN 0-900054-70-0.
- Beetle Larvae of the World. Entomological Society of America. ISBN 0-643-05506-1.
- Grimaldi, David; Michael S. Engel (2005). Evolution of the Insects. ISBN 0-521-82149-5.
- Harde, K. W. (1984). A Field Guide in Color to Beetles. pp. 7–24. ISBN 0-7064-1937-5.
- White, R. E. (1983). Beetles. Houghton Mifflin. ISBN 0-395-91089-7.
Ligazóns externas[editar | editar a fonte]
- Coleoptera Arquivado 09 de maio de 2012 en Wayback Machine. do Tree of Life Web Project
- (en alemán) Käfer der Welt
- Atlas de coleópteros
- Escaravellos – Coleoptera
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
