
Alternancia de xeracións

A alternancia de xeracións (tamén chamada metaxénese) é un termo usado principalmente para describir os ciclos vitais de plantas (entendendo por tales aquí as Archaeplastida). Un individuo gametófito multicelular, que é haploide, é dicir, con n cromosomas, alterna no ciclo vital da especie cun esporófito multicelular, que é diploide, é dicir, con 2n cromosomas (n pares). Un esporófito maduro produce esporas por meiose, un proceso que reduce o número de cromosoma á metade, de 2n a n. Como a meiose é un paso clave na alternancia de xeracións, é probable que teña unha función adaptativa fundamental. A natureza desta función aínda non se coñece ben, pero as dúas principais ideas que se esgrimen son que a meiose é adaptativa porque facilita a reparación dos danos no ADN ou que xera variabilidade xenética.
As esporas haploides xerminan e crecen por mitose orixinando gametófitos haploides. Na madurez, o gametófito produce gametos por mitose, o cal non altera o seu número de cromosomas. Dous gametos (orixinados en dous organismos de diferentes especies ou da mesma especie) fusiónanse para producir un cigoto, o cal se desenvolve dando un esporófito diploide. Este ciclo, do gametófito ao esporófito (ou igualmente do esporófito ao esporófito), é o modo no cal as plantas terestres e moitas algas realizan a súa reprodución sexual.
As relacións entre o esporófito e o gametófito varían entre diferentes grupos de plantas. Nas algas que teñen alternancia de xeracións, o esporófito e o gametófito son organismos separados independentes, os cales poden ter aparencia similar ou non. En hepáticas, musgos e antocerotas, o esporófito está menos desenvolvido que o gametófito e é en gran medida dependente del. Aínda que os esporófitos de musgos e antocerotas poden fotosintetizar, requiren un fotosintato adicional do gametófito para manter o crecemento e desenvolvemento das esporas e dependen del para a obtención de auga, nutrientes minerais e nitróxeno.[1][2] Ao contrario, en todas as plantas vasculares modernas o gametófito está menos desenvolvido que o esporófito, aínda que os seus antepasados do Devoniano tiñan gametófitos e esporófitos de complexidade aproximadamente equivalente.[3] En fentos o gametófito é un pequeno e aplanado prototalo autótrofo sobre o cal o esporófito novo é brevemente dependente para a súa nutrición, pero axiña crece e faise independente. En plantas con flor, a redución do gametófito é moito máis extremo, xa que consta só dunhas poucas células que crecen enteiramente dentro do esporófito.
Todos os animais desenvólvense de forma diferente a esta. Un animal maduro é diploide e, por tanto, en certo sentido, é equivalente ao esporófito das plantas. Porén, un animal produce directamente gametos haploides por meiose. Non producen ningún tipo de esporas haploides con capacidade de dividirse, polo que nunca se orixina nada similar a un gametófito haploide. Non hai alternancia entre formas haploides e diploides. Ás veces fálase de alternancia de xeracións en animais que teñen distintas fases no seu ciclo vital (como a fase polipoide e medusoide dos cnidarios), pero esta non é unha verdadeira alternancia equiparable á das plantas.
Os ciclos de vida das plantas e algas con estadios alternantes haploides e diploides multicellares denomínanse diplohaplontes (ou haplodiplontes, diplobiontes ou dibiónticos). Os ciclos de vida, como os dos animais, nos cales hai un só estadio multicelular diploide son denominados diplontes. (Os ciclos vitais nos cales hai só un estadio multicellar haploide denomínanse haplontes.)
Definición[editar | editar a fonte]
A discusión da 'alternancia de xeracións' de arriba considedra a alternancia dunha forma diploide multicelular cunha forma haploide multicelular como característica definitoria, inependentemente de se estas formas son de vida libre ou non.[4] Nalgunhas especies, como a alga Ulva lactuca, as formas diploide e haploide son ambas organismos de vida libre independentes, esencialmente idénticos en aparencia e por tanto denominados isomórficos. Os gametos haploides que nadan libremente forman un cigoto diploide, que xermina orixinando un esporófito diploide multicelular. O esporófito produce por meiose esporas haploides que nadan libres, que xerminan dando gmetófitos haploides.[5]
Porén, noutros grupos, tanto o esporófito coma o gametófito están moi reducidos e son incapaces de levar unha vida libre. Por exemplo, en todas as briófitas a xeración gametofítica é a dominante e o esporófito depende do gametófito. Ao contrario, en todas as plantas vasculares modernas os gametófitos están enormemente reducidos, aínda que a evidencia fósil nos di que derivaron de devanceiros isomórficos.[3] Nas espermatófitas, o gametófito feminino desenvólvese totalmente dentro do esporófito, o cal o protexe e nutre e tamén ao embrión esporofítico que se produce. Os grans de pole, que son os gametófitos masculinos, están reducidos a só unhas poucas células (en moitos casos a só tres células). Aquí a noción de dúas xeracións é menos obvia; como Bateman & Dimichele dixeron "os esporófitos e gametófitos funcionan de feito como un só organismo".[6] O termo alternativo 'alternancia de fases' pode entón ser máis apropiado.[7]
Historia[editar | editar a fonte]
Os debates sobre a alternancia de xeracións a inicios do século XX foron ás veces confusos porque coexistían varias maneiras de clasificar as "xeracións" (sexual fronte asexual, gametófito fronte a esporófito, haploide fronte a diploide etc.).[8]
Inicialmente, Chamisso e Steenstrup describiron a sucesión de xeracións organizadas de forma diferente (sexual e asexual) en animais como "alternancia de xeracións", cando estudaban o desenvolvemento de tunicados, cnidarios e trematodos.[8] Este fenómeno tamén se coñece como heterogamia. Actualmente, o termo "alternancia de xeracións" está asociado case exclusivamente cos ciclos de vida das plantas, especificamente coa alternancia entre gametófitos haploides e esporófitos diploides.[8]
Wilhelm Hofmeister demostrou a alternancia morfolóxica de xeracións en plantas,[9] entre unha xeración que produce esporas (esporófito) e unha xeración que produce gametos (gametófito).[10][11] Nesa época xurdiu un debate centrado na orixe da xeración asexual das plantas terrestres (é dicir, o esporófito) que se considera un conflito entre as teorías da alternancia de xeracións antitética (Čelakovský, 1874) e homóloga (Pringsheim, 1876).[8] Čelakovský usou as palabras esporófito e gametófito.
Eduard Strasburger (1874) descubriu a alternancia entre as fases nucleares diploide e haploide,[8] tamén chamada alternancia citolóxica de fases nucleares.[12] Aínda que moi a miúdo coinciden, a alternancia morfolóxica e a das fases nucleares son ás veces independentes entre si, por exemplo, en moitas algas vermellas, a mesma fase nuclear pode corresponder a dúas distintas xeracións morfolóxicas.[12] Nalgúns fentos que perderon a reprodución sexual, non hai cambio de fase nuclear, pero mantense a alternancia de xeracións.[13]
Alternancia de xeracións en plantas[editar | editar a fonte]
Elementos fundamentais[editar | editar a fonte]
O diagrama do principio deste artigo mostra os elementos fundamentais da alternancia de xeracións en plantas. Nos diferentes grupos de plantas hai moitas variacións deste esquema básico, que se describirán máis adiante. Empezando pola parte dereita do diagrama, os procesos indicados son os seguintes:[14]
- Dous gametos haploides unicelulares, cada un dos cales contén n cromosomas que non forman pares, fusiónanse para formar un cigoto unicelular diploide, que agora contén 2n cromosomas en total, distrubuídos en n pares.
- O cigoto diploide unicelular xermina, dividíndose polo proceso normal de división, a mitose, a cal mantén o número de cromosomas en 2n. O resultado é un organismo diploide multicelular, chamado esporófito (porque na madurez produce esporas).
- Cando chega á madurez, o esporófito produce un ou máis esporanxios, que son os órganos que producen as células nais das esporas diploides (esporocitos). Nestes prodúcense divisións polo proceso de división especial da meiose, que reduce o número de cromosomas á metade. Isto orixina inicialmente catro esporas haploides unicelulares, cada unha das cales contén n cromosomas que non forman pares.
- As esporas haploides unicelulares xerminan, dividíndose por mitose, polo que o número de cromosomas das células fillas se mantén en n. O resultado é un organismo haploide multicelular, chamado gametófito (porque pode producir gametos).
- Cando chega á madurez, o gametófito produce un ou máis gametanxios, que son os órganos que producen gametos haploides. Polo menos un dos tipos de gameto posúe algún mecanismo para buscar o outro gameto e chegar ata el para fusionarse con el.
A 'alternancia de xeracións' no ciclo vital establécese entre unha xeración diploide (2n) de esporófitos e outra haploide (n) de gametófitos.

A situación é bastante diferente á que se observa en animais, onde o proceso fundamental é que un individuo diploide (2n) produce directamente gametos haploides (n) por meiose. Non se producen esporas (é dicir, células haploides capaces de realizar mitoses), polo que nunca se forma un individuo multicelular haploide. Os gametos unicelulares son as únicas entidades que son haploides.
Variacións[editar | editar a fonte]
O diagrama mostrado arriba é unha boa representación do ciclo vital dalgunhas algas multicelulares (por exemplo, as do xénero Cladophora), que teñen esporófitos e gametófitos de aparencia case idéntica e que non teñen diferentes tipos de esporas ou gametos.[15]
Porén, hai moitas posibles variacións dos elementos fundamentais dun ciclo vital con alternancia de xeracións. Cada variación pode ocorrer separadamene ou en combinación, o que ten como resultado unha enorme variedade de ciclos de vida. Os termos usados polos botánicos para describir estes ciclos son tamén enormemente variados. Como dixeron Bateman e Dimichele: "[...] a alternancia de xeracións converteuse nun pantano terminolóxico; a miúdo un termo representa varios conceptos ou un concepto está representado por varios termos."[16]
Posibles variacións son:
- Importancia relativa do eporófito e o gametófito.
- Igual importancia (homomorfia ou isomorfia).
As algas filamentosas do xénero Cladophora, que se encontran predominantemente en augas doces, teñen esporófitos diploides e gametófitos haploides que son externamente indistinguibles.[17] Ningunha planta terrestre viva ten esporófitos e gametófitos con igual dominancia, aínda que algunhas teorías da evolución da alternancia de xeracións suxiren que as plantas terrestres ancestrais si tiñan. - Distinta importancia (heteromorfia ou anisomorfia).

Gametófito do musgo Mnium hornum. - Gametófito dominante (ciclo gametofítico).
En musgos, hepáticas e antocerotas, a forma dominante é o gametófito haploide. O esporófito diploide non pode ter unha existencia independente, e obtén a maior parte da súa nutrición do gametófito parental, e non ten clorofila cando madura.[18]
Esporófito do fento Blechnum discolor. - Esporófito dominante (ciclo esporofítico).
En fentos, tanto o esporófito coma o gametófito poden vivir independentemente, pero a forma dominante é o esporófito diploide. O gametófito haploide é moito máis pequeno e simple en estrutura. Nas plantas con semente, o gametófito está aínda máis reducido (no caso mínimo a só tres células), e obtén toda a súa nutrición do esporófito. A redución tan extrema de tamaño do gametófito e a súa retención dentro do esporófito significa que aplicar ás plantas con sementes o termo 'alternancia de xeracións' é un tanto enganoso: "os esporófitos e gametófitos funcionan de feito como un só organismo".[6] Por iso algúns autores prefiren neste caso falar de 'alternancia de fases'.[7]
- Gametófito dominante (ciclo gametofítico).
- Igual importancia (homomorfia ou isomorfia).
- Diferenciación dos gametos.
- Ambos os gametos son iguais (isogamia).
Como outras especies de Cladophora, a especie C. callicoma ten gametos flaxelados que son idénticos en aspecto e na capacidade e moverse.[17] - Gametos de dous tamaños distintos (anisogamia).
- Ambos de motilidade similar.
Especies de Ulva, a leituga de mar, teñen gametos móbiles con dous flaxelos. Porén son de dous tamaños: gametos 'femininos' máis grandes e 'masculinos' máis pequenos.[19] - Un gameto grande e sésil e outro pequeno e móbil (oogamia). Os megagametos máis grandes e sésiles son os ovos (ovocélula, oogonia) e os microgametos móbiles pequenos son os espermatozoides (ou anterozoides). O grao de mobilidade do espermatozoide pode ser moi limitado (como nas plantas con flor) pero sempre teñen algún modo de moverse cara aos ovos sésiles. Cando os espermatozoides e os ovos se producen en diferentes tipos de gametanxios (que é o que ocorre case sempre), os que producen os espermatozoides denomínanse anteridios e os que producen os ovos arquegonios.

Gametófito de Pellia epiphylla con esporófitos crecendo a partir dos restos do arquegonio. - Anteridios e arquegonios no mesmo gametófito, o cal se denomina monoico. (Moitas fontes, incluíndo as que tratan dos briófitos, usan o termo 'monoecio' para esta situación e 'diecio' para a contraria.[20][21] Aquí 'monoecio' e 'diecio' utilízanse só para esporófitos.)
Na hepática Pellia epiphylla o gametófito é a xeración dominante. É monoico: os pequenos anteridios avermellados produtores de espermatozoides están espallados por toda a parte central, mentres que os arquegonios produtores de ovos son máis periféricos.[22] - Anteridios e arquegonios en distintos gametófitos, que se denominan dioicos.
O musgo Mnium hornum ten o gametófito como xeración dominante. É dioico: as plantas macho producen só anteridios en rosetas terminais; as plantas femininas producen só arquegonios en forma de cápsulas con taliño.[23] Os gametófitos de plantas con sementes son tamén dioicos. Porén, o esporófito parental pode ser monoecio, producindo no mesmo individuo tanto os gametófitos masculinos coma os femininos, ou diecio, producindo gametófitos dun só xénero (gametófitos masculinos ou femininos en individuos separados). Os gametófitos das plantas con sementes están extremadamente reducidos en tamaño; o arquegonio consta só dun pequeno número de células, e o gametófito masculino completo pode constar só de dúas ou tres células.[24]
- Anteridios e arquegonios no mesmo gametófito, o cal se denomina monoico. (Moitas fontes, incluíndo as que tratan dos briófitos, usan o termo 'monoecio' para esta situación e 'diecio' para a contraria.[20][21] Aquí 'monoecio' e 'diecio' utilízanse só para esporófitos.)
- Ambos de motilidade similar.
- Ambos os gametos son iguais (isogamia).
- Diferenciación das esporas.
- Esporas do mesmo tamaño (homosporia ou isosporia).
Especies de Equisetum teñen esporas do mesmo tamaño.[25] - Esporas de dous tamaños distintos ('heterosporia ou anisosporia): megasporas maiores e microsporas máis pequenas. Cando os dous tipos de esporas se producen en diferentes tipos de esporanxios, estes denomínanse megasporanxios e microsporanxios. Unha megaspora a miúdo (pero non sempre) desenvólvese a expensas das outras tres células orixinadas na meiose, as cales abortan.
- Megasporanxios e microsporanxios no mesmo esporófito, que se denomina monoecio.
A maioría das plantas con flor están dentro desta categoría. Así, a flor dunha azucena é hermafrodita, xa que contén seis estames (os microsporanxios) que producen microsporas, que se desenvolven en grans de pole (os microgametófitos), e tres carpelos fusionados, que producen megasporanxios tegumentados (óvulos ou primordios seminais) cada un dos cales produce unha megaspora, que se desenvolve dentro do megasporanxio para producir o megagametófito. Noutras plantas, como a abeleira, algunhas flores teñen só estames (flores unisexuais masculinas) e outras só carpelos (flores unisexuais femininas), pero a mesma planta (é dicir, o esporófito) ten ambos os tipos de flores, polo que é monoecia.
Flores de acivro, unha especie dioica: a planta macho arriba e a femia abaixo (cortáronse follas para que se vexan mellor as flores). - Megasporanxios e microsporanxios en diferentes esporófitos, que se denominan diecios.
Unha planta de acivro (Ilex aquifolium) produce tanto flores 'masculinas' que teñen só estames (microsporanxios) funcionais, que producen microsporas, que se desenvolven en grans de pole (microgametófitos) coma flores 'femininas', que teñen só carpelos funcionais, que producen megasporanxios tegumentados (óvulos ou primordios seminais), que conteñen unha megaspora, que se desenvolve dando un megagametófito multicelular.
- Megasporanxios e microsporanxios no mesmo esporófito, que se denomina monoecio.
- Esporas do mesmo tamaño (homosporia ou isosporia).




(Nota: Non se deben confundir os óvulos dos animais, que son unha soa célula (o gameto feminino), cos óvulos ou primordios seminais das plantas, que son unha estrutura multicelular (macrosporanxio), onde se forma a magaspora).
Nas plantas hai algunhas correlacións entre estas variacións, pero son só correlacións e non absolutas. Por exemplo, nas plantas con flor, as microsporas producen finalmente microgametos (espermatozoides, núcleos do pole) e as megasporas producen finalmente megagametos (ovos, ovocélulas). Porén, en fentos e grupos relacionados hai grupos con esporas indiferenciadas pero gametófitos diferenciados. Por exemplo, o fento Ceratopteris thalictrioides ten esporas dun único tipo, que varían en tamaño de forma continua. As esporas menores tenden a xerminar en gametófitos que producen só anteridios produtores de espermatozoides.[25]
Un ciclo de vida complexo[editar | editar a fonte]

O diagrama mostra a alternancia de xeracións nunha especie heteromófica, esporofítica, oogamética, dioica, heterospórica e diecia. Un exemplo de planta con sementes pode ser un salgueiro (a maioría das especies do xénero Salix son diecias).[26] Comezando desde o centro do diagrama, os procesos implicados son:
- Un ovo inmóbil, contido no arquegonio, fusiónase cun espermatozoide móbil, liberado dun anteridio. O cigoto resultante pode ser 'macho' ou 'femia'.
- Un cigoto 'macho' desenvólvese por mitose orixinando un microsporófito, o cal na madurez produce máis microsporanxios. As microsporas desenvólvense dentro do microsporanxio por meiose.
Nun salgueiro (como en todas as plantas con sementes) o cigoto desenvólvese primeiro dando un embrión microsporófito dentro do óvulo ou primoridio seminal (un megasporanxio encerrado nunha ou máis capas protectoras de tecido coñecidas como tegumento). Na madurez, estas estruturas convértense en sementes. Posteriormente a semente libérase, xermina e crece dando lugar a unha árbore madura. Un salgueiro 'macho' (un microsporófito) produce flores que só teñen estames, as anteras dos cales son os microsporanxios. - As microsporas xerminan producindo microgametófitos; na madurez prodúcense anteridios. Os espermatozoides desenvólvense dentro dos anteridios.
Nun salgueiro, as microsporas non son liberadas da antera (o microsporanxio), pero desenvólvense orixinando no seu interior grans de pole (microgametófitos). O gran de pole completo desprázase (por exemplo levado polo vento ou insectos) ata a parte feminina e crécelle un tubo polínico que chega ao óvulo (megagametófito). Dentro do gran de pole está o espermatozoide (núcleos celulares) que baixa polo tubo polínico e chega ao ovo situado no interior do óvulo ou primorido seminal. - Un cigoto 'femia' desenvólvese por mitose nun megasporófito, que na súa madurez produce megasporanxios. As megasporas desenvólvense dentro do megasporanxio; xeralmente unha das catro esporas producidas por meiose faise máis grande a expensas das outras tres, que desaparecen.
Os salgueiros 'femias' (megasporófitos) producen flores que só teñen carpelos (follas modificadas que levan os megasporanxios). - As megasporas xerminan producindo megagametófitos; na madurez prodúcense arquegonios. Os ovos desenvólvense dentro dos arquegonios.
Os carpelos dun salgueiro producen óvulos, megasporanxios envoltos en tegumentos. Dentro de cada óvulo, unha megaspora desenvólvese por mitose nun megagametófito. Un arquegonio desenvólvese dentro do megagametófito e produce un ovo. A 'xeración' gametofítica permanece protexido no interior do esporófito excepto os graos de pole (reducidos a tres células contidas dentro da parede da microspora).
- Un cigoto 'macho' desenvólvese por mitose orixinando un microsporófito, o cal na madurez produce máis microsporanxios. As microsporas desenvólvense dentro do microsporanxio por meiose.
Ciclos de vida de diferentes grupos de plantas[editar | editar a fonte]
O termo "plantas" utilízase aquí co significado de Archaeplastida, é dicir glaucófitas, algas vermellas, algas verdes e plantas terrestres.
A alternancia de xeracións ocorre en case todas as algas verdes e vermellas multicelulares, tanto en formas de auga doce (como Cladophora) coma mariñas (como Ulva). Na maioría, as xeracións son homomórficas (isomórficas) e de vida libre. Algunhas especies de algas vermellas teñen unha alternancia de xeracións trifásica, na cal hai unha fase gametofítica e dúas fases esporofíticas distintas.
As plantas terrestres teñen todas alternancia de xeracións heteromórfica (anisomórfica), na cal o esporófito e o gametófito son claramente diferentes. Todas as briófitas sensu lato, é dicir hepáticas, musgos e antocerotas, teñen a xeración gametofítica máis desenvolvida que a esporofítica. Como ilustración, consideremos un musgo monoico. Na planta madura (o gametófito) desenvólvense anteridios e arquegonios. En presenza de auga que molla a planta, o anterozoide biflaxelado formado nos anteridios nada ata o arquegonio e alí ocorre a fecundación, dando lugar á produción dun esporófito diploide. O esporófito crece desde o arquegonio. O seu corpo está formado por un longo pedúnculo ou filamento que leva na parte superior unha cápsula dentro da cal están as células produtoras de esporas, que mediante meiose producen esporas haploides. A maioría dos musgos dependen do vento para a dispersión das esporas, aínda que Splachnum sphaericum é entomófilo, xa que os insectos dispersan as súas esporas.
-
 Diagrama da alternancia de xeracións en hepáticas.
Diagrama da alternancia de xeracións en hepáticas. -
 Diagrama do ciclo vital dun musgo.
Diagrama do ciclo vital dun musgo. -
 Diagrama do ciclo vital dun antocerota.
Diagrama do ciclo vital dun antocerota.



En fentos e grupos asociados como os licopodios e equisetos, a planta que se observa no campo é o esporófito diploide. As esporas haploides orixínanse en soros situados na parte inferior da fronde e son dispersadas polo vento (ou en certos casos flotando na auga). Se as condicións son as axeitadas, a espora xermina dando lugar a unha planta pouco visible chamada protalo. O protalo haploide non lembra o esporófito e, por tanto, estas plantas teñen unha alternancia de xeracións heteromórfica. O protalo ten curta vida, pero leva a cabo a reprodución sexual, producindo o cigoto diploide, que despois medra desde o protalo (o cal dexenera) formando o esporófito.
-
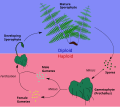
Diagrama da alternancia de xeracións en fentos.. -

Un gametófito (protalo) de Dicksonia sp. -

Un esporófito de Dicksonia antarctica. -

Parte inferior da fronde de Dicksonia antarctica mostrando os soros (estruturas formadoras de esporas).




Nas plantas con sementes (espermatófitas) o esporófito é a fase dominante multicelular, pero dunha maneira aínda máis esaxerada que nos fentos; os gametófitos están fortemente reducidos en tamaño e son moi diferentes en morfoloxía. A xeración gametofítica completa, coa soa excepción dos grans de pole (microgametófitos), está contida dentro do esporófito. O ciclo vital dunha planta con flores dioica anxiosperma, como o salgueiro, foi explicada con detalle nunha sección anterior. O ciclo de vida das ximnospermas é similar. Porén, as plantas con flores teñen ademais un fenómeno adicional chamado 'dobre fertilización'. Dous núcleos espermáticos (equivalentes a gametos masculinos) contidos no gran de pole (o microgametófito), entran no arquegonio do megagametófito; un fusiónase co núcleo da ovocélula (gameto feminino) para formar un cigoto, e o outro fusiónase con outros dous núcleos do gametófito para formar o 'endosperma', que é un tecido que nutre ao embrión en desenvolvemento.
Aparición na evolución da fase diploide dominante[editar | editar a fonte]
Propúxose que a base para explicar a aparición durante a evolución da fase diploide do ciclo de vida (esporófito) como fase dominante (por exemplo en plantas vasculares) é que a diploidía permite enmascarar a expresión de mutacións deletéreas por complementación xenética.[27][28] Así, se un dos xenomas parentais nas células diploides contén mutacións que orixinan defectos en produtos xénicos, estas deficiencias poderían ser compensadas polo outro xenoma parental (o cal, non obstante, pode ter os seus propios defectos noutros xenes). A medida que a fase diploide se foi facendo predominante, o efecto enmascarador probablemente permitía que o tamaño do xenoma e, por tanto, o contido de información, se incrementase sen a constrición de ter que mellorar a exactitude da replicación do ADN. A oportunidade de incrementar a información contida a un baixo custo era vantaxosa porque permitía que se codificasen novas adaptacións. Esta idea púxose en dúbida ao apareceren evidencias que mostraban que a selección non é máis efectiva na fase haploide que na diploide do ciclo de vida de musgos e anxiospermas.[29]
-
 Ciclo de vida das anxiospermas.
Ciclo de vida das anxiospermas. -
 Extremo do estame do tulipán mostrando os grans de pole (microgametófitos)
Extremo do estame do tulipán mostrando os grans de pole (microgametófitos) -
 Óvulos (megagametófitos) de plantas: óvulo de ximnosperma á esquerda, óvulo de anxiosperma (dentro do ovario) á dereita.
Óvulos (megagametófitos) de plantas: óvulo de ximnosperma á esquerda, óvulo de anxiosperma (dentro do ovario) á dereita. -
 Dobre fertilización.
Dobre fertilización.




Procesos similares noutros organismos[editar | editar a fonte]
Cromalveolados[editar | editar a fonte]
Algúns organismos actualmente clasificados nos Chromalveolata, que non son plantas no sentido usado aquí, mostran alternancia de xeracións. Os foraminíferos sofren unha alternancia de xeracións heteromórfica entre as formas gamontes haploides e formas agamontes diploides. O organismo haploide unicelular é tipicamente máis grande que o organismo diploide.
Fungos[editar | editar a fonte]
Os micelios dos fungos son tipicamente haploides. Cando os micelios de diferentes tipos de apareamento se encontran, producen dúas células con forma de bóla multinucleadas, as cales se unen por medio de "pontes de apareamento". Os núcleos móvense dun micelio a outro, formando un heterocarion (que significa "núcleos diferentes"). Este proceso denomínase plasmogamia. A fusión real para formar núcleos diploides denomínase cariogamia, e non pode ocorrer ata que se forman os esporanxios. A cariogamia produce un cigoto diploide, que é un esporófito de curta vida que axiña sofre a meiose para formar esporas haploides. Cando as esporas xerminan, orixinan novos micelios.
Mofos mucosos[editar | editar a fonte]
O ciclo de vida dos mofos mucosos é moi similar ao dos fngos. As esporas haploides xerminan formando células que se enxamian ou mixamebas. Estas fusiónanse nun proceso denominado plasmogamia e cariogamia para formar un cigoto diploide. O cigoto desenvólvese orixinando un plasmodio, e o plasmodio maduro produce, dependendo das especies, corpos frutíferos que conteñen esporas haploides.
Animais[editar | editar a fonte]
A alternancia entre unha xeración multicelular diploide e outra multicelular haploide nunca se encontra nos animais.[30] Nalgúns animais, hai unha alternancia entre fases partenoxenéticas e de reprodución sexual (heterogamia) ou fases sexuais e asexuais. Ambas as fases son diploides. Isto foi ás veces chamado "alternancia de xeracións",[31] pero non é en absoluto comparable coa das plantas.
Notas[editar | editar a fonte]
- ↑ Thomas, R.J.; Stanton, D.S.; Longendorfer, D.H. & Farr, M.E. (1978). "Physiological evaluation of the nutritional autonomy of a hornwort sporophyte". Botanical Gazette 139 (3): 306–311. doi:10.1086/337006.
- ↑ Glime, J.M. (2007). Bryophyte Ecology: Vol. 1 Physiological Ecology (PDF) (Informe). Michigan Technological University and the International Association of Bryologists. Consultado o 2013-03-04.
- ↑ 3,0 3,1 Kerp, H.; Trewin, N.H. & Hass, H. (2003). "New gametophytes from the Lower Devonian Rhynie Chert". Transactions of the Royal Society of Edinburgh: Earth Sciences 94 (4): 411–428. doi:10.1017/S026359330000078X.
- ↑ Taylor, Kerp & Hass 2005
- ↑ ""Plant Science 4 U". Consultado o 5 xullo de 2016.
- ↑ 6,0 6,1 Bateman & Dimichele 1994, p. 403
- ↑ 7,0 7,1 Stewart & Rothwell 1993
- ↑ 8,0 8,1 8,2 8,3 8,4 Haig, David (2008). "Homologous versus antithetic alternation of generations and the origin of sporophytes" (PDF). The Botanical Review 74 (3): 395–418. doi:10.1007/s12229-008-9012-x. Consultado o 2014-08-17.
- ↑ Svedelius, Nils (1927). "Alternation of Generations in Relation to Reduction Division". Botanical Gazette 83 (4): 362–384. JSTOR 2470766. doi:10.1086/333745.
- ↑ Hofmeister, W. (1851). Vergleichende Untersuchungen der Keimung, Entfaltung und Fruchtbildildiung höherer Kryptogamen (Moose, Farne, Equisetaceen, Rhizocarpeen und Lycopodiaceen) und der Samenbildung der Coniferen (en alemán). Leipzig: F. Hofmeister. Consultado o 2014-08-17.. Hai tamén unha tradución en inglés: Currey, Frederick (1862). On the germination, development, and fructification of the higher Cryptogamia, and on the fructification of the Coniferæ. London: Robert Hardwicke. Consultado o 2014-08-17.
- ↑ Feldmann, J. & Feldmann, G. (1942). "Recherches sur les Bonnemaisoniacées et leur alternance de generations" (PDF). Ann. Sci. Natl. Bot., ser. 11 (en French) 3: 75–175. Arquivado dende o orixinal (PDF) o 19 de agosto de 2014. Consultado o 09 de novembro de 2016., p. 157
- ↑ 12,0 12,1 Feldmann, J. (1972). "Les problèmes actuels de l'alternance de génerations chez les Algues". Bulletin de la Société Botanique de France (en French) 119: 7–38. doi:10.1080/00378941.1972.10839073.
- ↑ Schopfer, P.; Mohr, H. (1995). "Physiology of Development". Plant physiology. Berlin: Springer. pp. 288–291. ISBN 3-540-58016-6.
- ↑ A non ser que se indique outra cousa, o texto de toda esta sección está baseado en Foster & Gifford 1974, Sporne 1974a e Sporne 1974b.
- ↑ Guiry & Guiry 2008
- ↑ Bateman & Dimichele 1994, p. 347
- ↑ 17,0 17,1 Shyam 1980
- ↑ Watson 1981, p. 2
- ↑ Kirby 2001
- ↑ Watson 1981, p. 33
- ↑ Bell & Hemsley 2000, p. 104
- ↑ Watson 1981, pp. 425–6
- ↑ Watson 1981, pp. 287–8
- ↑ Sporne 1974a, pp. 17–21.
- ↑ 25,0 25,1 Bateman & Dimichele 1994, pp. 350–1
- ↑ "Willows". Encyclopædia Britannica XIX (11th ed.). New York: Encyclopædia Britannica. 1911. Consultado o 2011-01-01.
- ↑ Bernstein, H.; Byers, G.S. & Michod, R.E. (1981). "Evolution of sexual reproduction: Importance of DNA repair, complementation, and variation". The American Naturalist 117 (4): 537–549. doi:10.1086/283734.
- ↑ Michod, R.E. & Gayley, T.W. (1992). "Masking of mutations and the evolution of sex". The American Naturalist 139 (4): 706–734. doi:10.1086/285354.
- ↑ Szövényi, Péter; Ricca, Mariana; Hock, Zsófia; Shaw, Jonathan A.; Shimizu, Kentaro K. & Wagner, Andreas (2013). "Selection is no more efficient in haploid than in diploid life stages of an angiosperm and a moss". Molecular Biology and Evolution 30 (8): 1929. PMID 23686659. doi:10.1093/molbev/mst095.
- ↑ Barnes et al. 2001, p. 321
- ↑ Scott 1996, p. 35
Véxase tamén[editar | editar a fonte]
Outros artigos[editar | editar a fonte]
Bibliografía[editar | editar a fonte]
- Barnes, R.S.K.; Calow, P.; Olive, P.J.W.; Golding, D.W. & Spicer, J.I. (2001). The Invertebrates: a synthesis. Oxford; Malden, MA: Blackwell. ISBN 978-0-632-04761-1.
- Bateman, R.M. & Dimichele, W.A. (1994). "Heterospory – the most iterative key innovation in the evolutionary history of the plant kingdom" (PDF). Biological Reviews of the Cambridge Philosophical Society 69: 315–417. Arquivado dende o orixinal (PDF) o 15 de abril de 2012. Consultado o 2010-12-30.
- Bell, P.R. & Hemsley, A.R. (2000). Green Plants: their Origin and Diversity (2nd ed.). Cambridge etc.: Cambridge University Press. ISBN 978-0-521-64109-8.
- Foster, A.S. & Gifford, E.M. (1974). Comparative Morphology of Vascular Plants (2nd ed.). San Francisco: W.H. Freeman. ISBN 978-0-7167-0712-7.
- Guiry, M.D.; Guiry, G.M. (2008). "Cladophora". AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. Consultado o 2011-07-21.
- Kirby, A. (2001). "Ulva, the sea lettuce". Monterey Bay Aquarium Research Institute. Arquivado dende o orixinal o 16 de maio de 2011. Consultado o 2011-01-01.
- Scott, Thomas (1996). Concise Encyclopedia Biology. Berlin: Walter de Gruyter. ISBN 978-3-11-010661-9.
- Shyam, R. (1980). "On the life-cycle, cytology and taxonomy of Cladophora callicoma from India". American Journal of Botany 67 (5): 619–24. JSTOR 2442655. doi:10.2307/2442655.
- Sporne, K.R. (1974a). The Morphology of Angiosperms. London: Hutchinson. ISBN 978-0-09-120611-6.
- Sporne, K.R. (1974b). The Morphology of Gymnosperms (2nd ed.). London: Hutchinson. ISBN 978-0-09-077152-3.
- Stewart, W.N. & Rothwell, G.W. (1993). Paleobotany and the Evolution of Plants (2nd ed.). Cambridge, UK: Cambridge University Press. ISBN 978-0-521-38294-6.
- Watson, E.V. (1981). British Mosses and Liverworts (3rd ed.). Cambridge, UK: Cambridge University Press. ISBN 978-0-521-28536-0.
- Taylor, T.N.; Kerp, H. & Hass, H. (2005). "Life history biology of early land plants: Deciphering the gametophyte phase". Proceedings of the National Academy of Sciences of the United States of America 102 (16): 5892–5897. PMC 556298. PMID 15809414. doi:10.1073/pnas.0501985102.